

DUB3 Deubiquitylating Enzymes Regulate Hippo Pathway Activity by Regulating the Stability of ITCH, LATS and AMOT Proteins
- PMID: 28061504
- PMCID: PMC5218808
- DOI: 10.1371/journal.pone.0169587
DUB3 Deubiquitylating Enzymes Regulate Hippo Pathway Activity by Regulating the Stability of ITCH, LATS and AMOT Proteins
Abstract
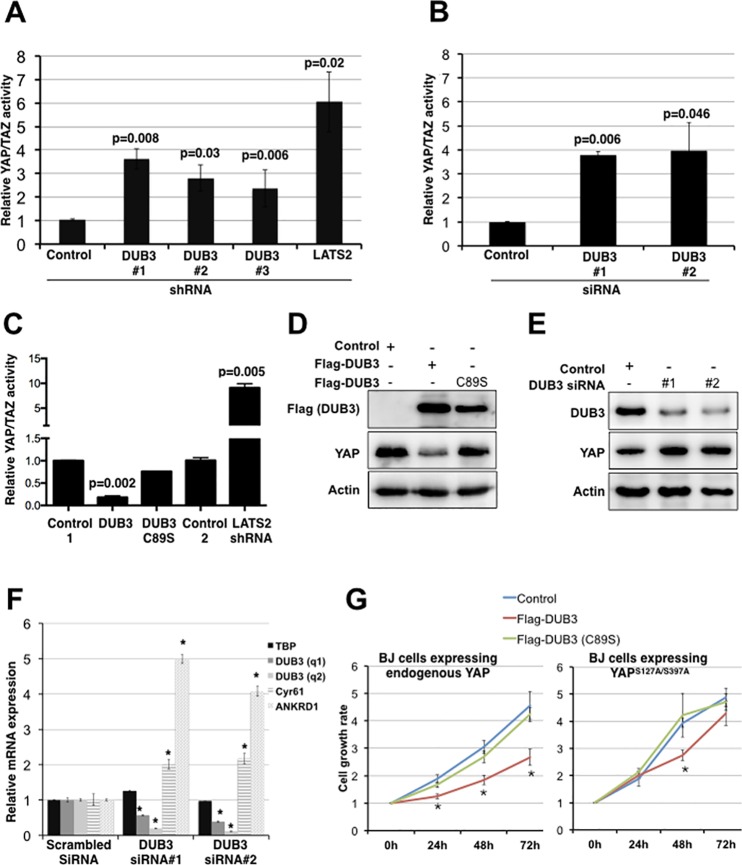
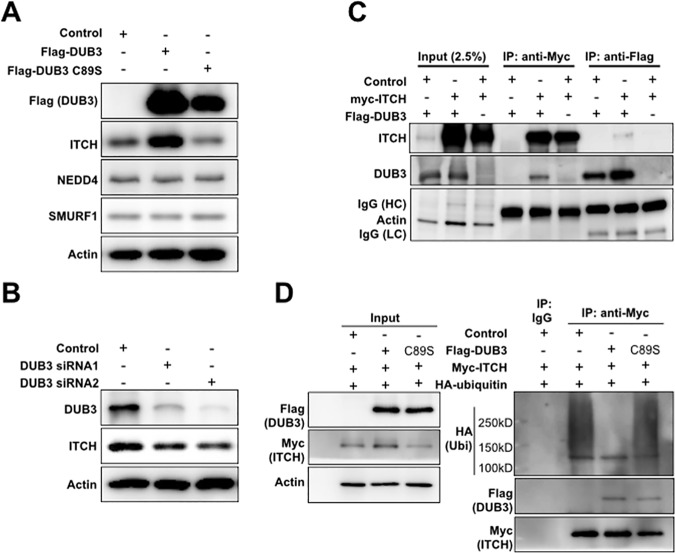
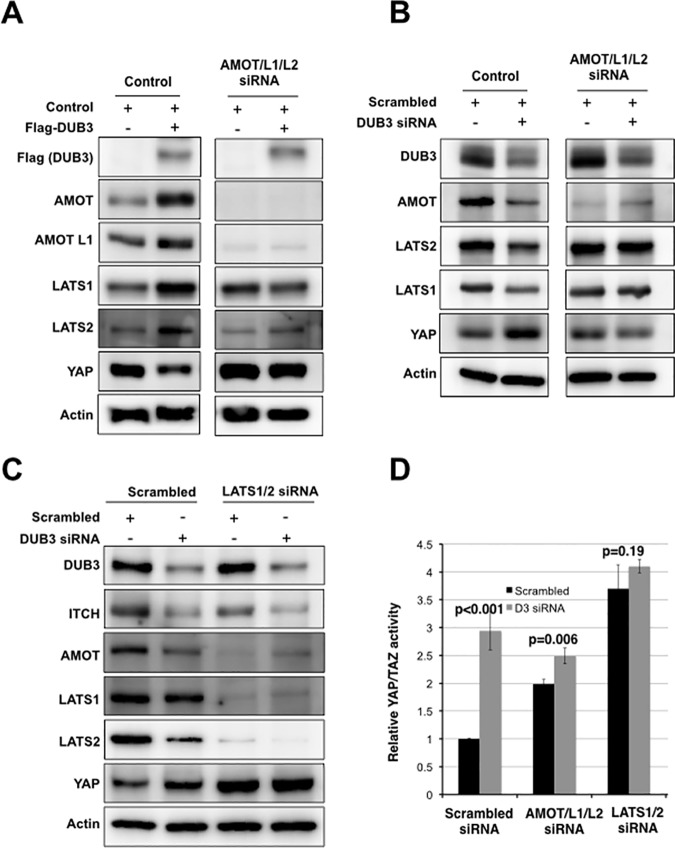
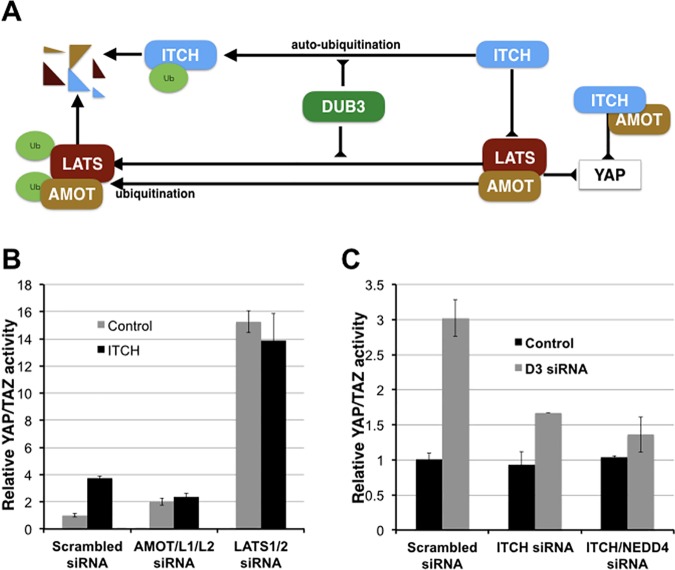
The YAP and TAZ transcriptional coactivators promote oncogenic transformation. Elevated YAP/TAZ activity has been documented in human tumors. YAP and TAZ are negatively regulated by the Hippo tumor suppressor pathway. The activity and stability of several Hippo pathway components, including YAP/TAZ, is regulated by ubiquitin mediated protein turnover and several ubiquitin ligase complexes have been implicated in human cancer. However, little is known about the deubiquitylating enzymes that counteract these ubiquitin ligases in regulation of the Hippo pathway. Here we identify the DUB3 family deubiquitylating enzymes as regulators of Hippo pathway activity. We provide evidence that DUB3 proteins regulate YAP/TAZ activity by controlling the stability of the E3 ligase ITCH, the LATS kinases and the AMOT family proteins. As a novel Hippo pathway regulator, DUB3 has the potential to act a tumor suppressor by limiting YAP activity.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Harvey K. F., Zhang X., and Thomas D. M. (2013) The Hippo pathway and human cancer. Nature reviews. Cancer 13, 246–257 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
