

Subangstrom Measurements of Enzyme Function Using a Biological Nanopore, SPRNT
- PMID: 28062043
- PMCID: PMC6044436
- DOI: 10.1016/bs.mie.2016.09.038
Subangstrom Measurements of Enzyme Function Using a Biological Nanopore, SPRNT
Abstract
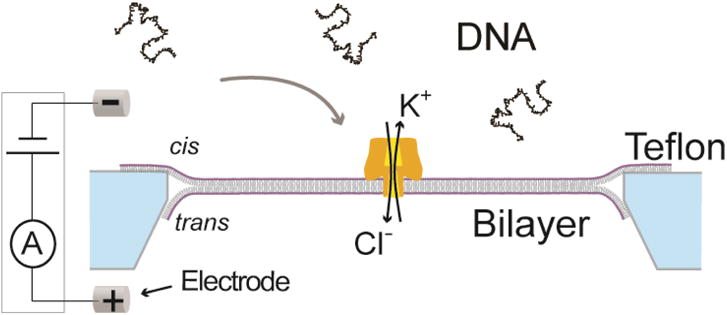
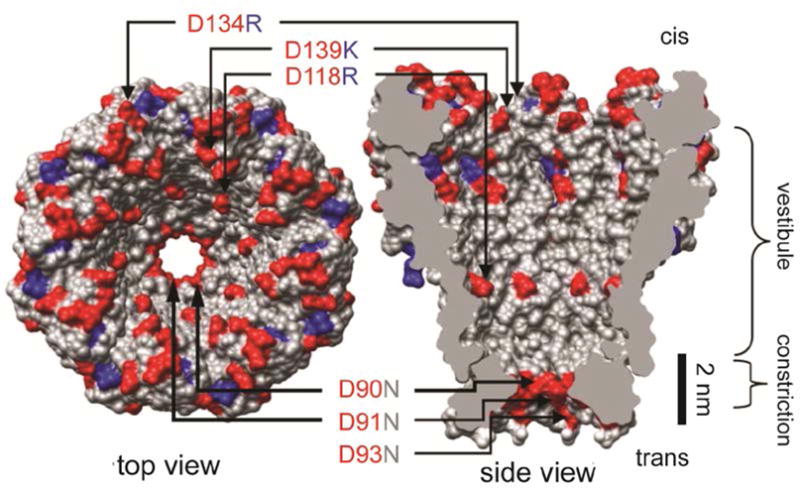
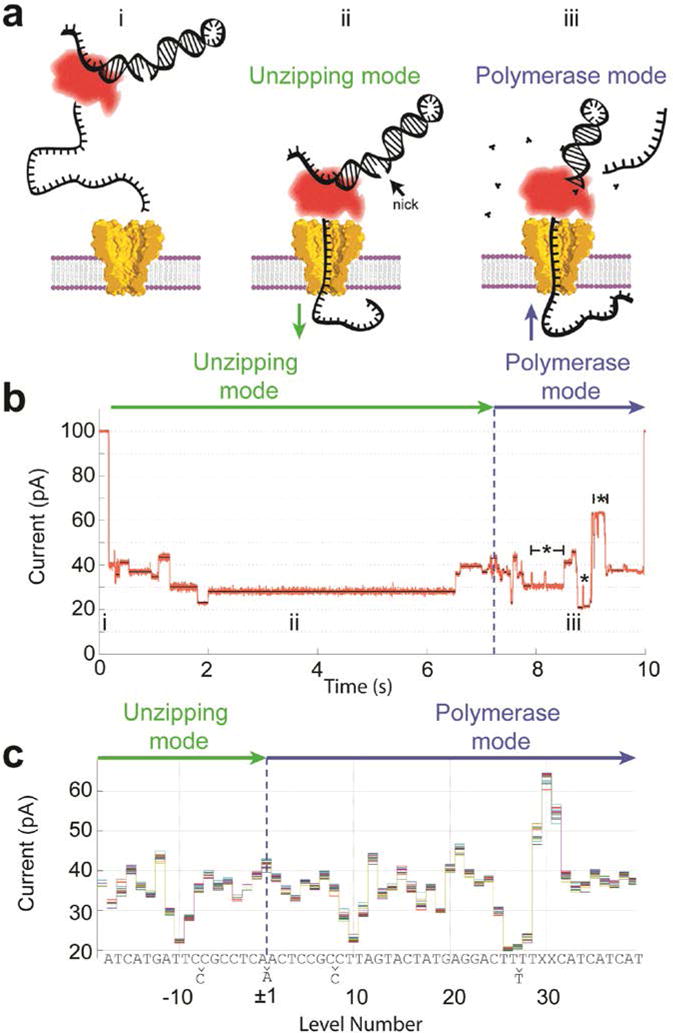
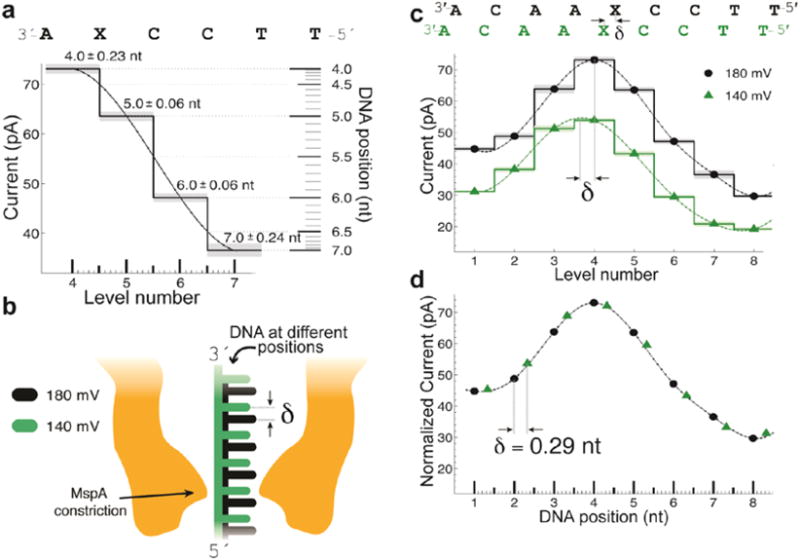
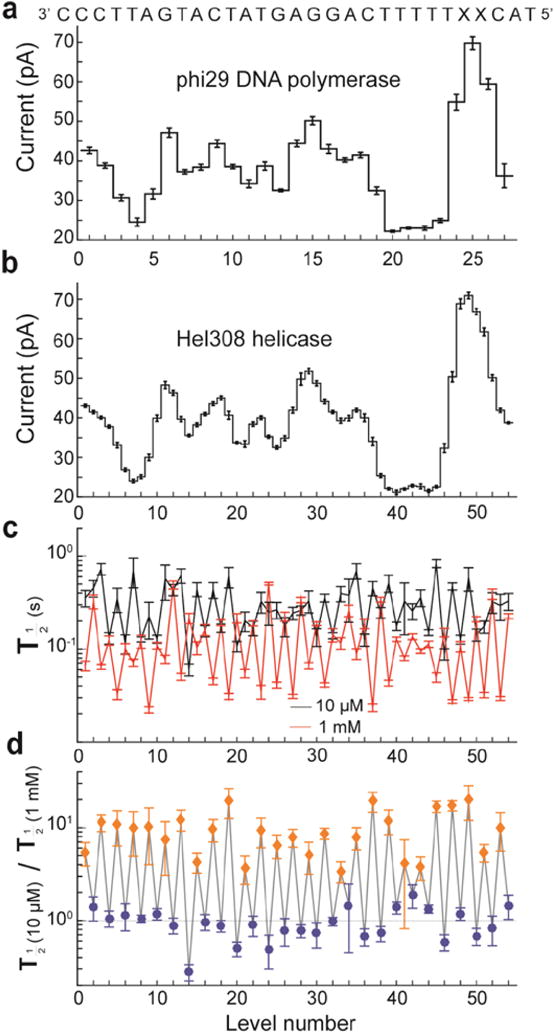
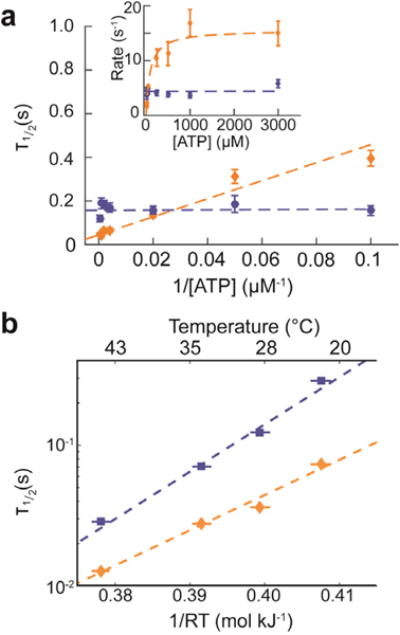
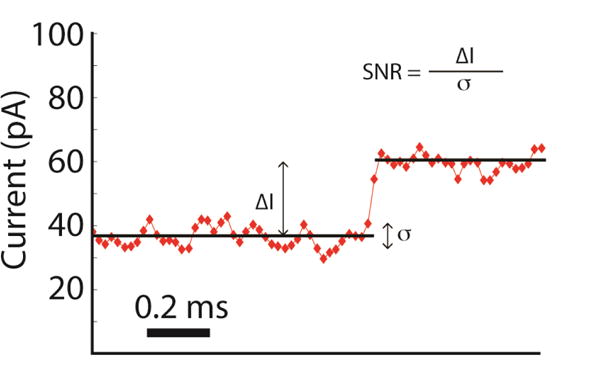
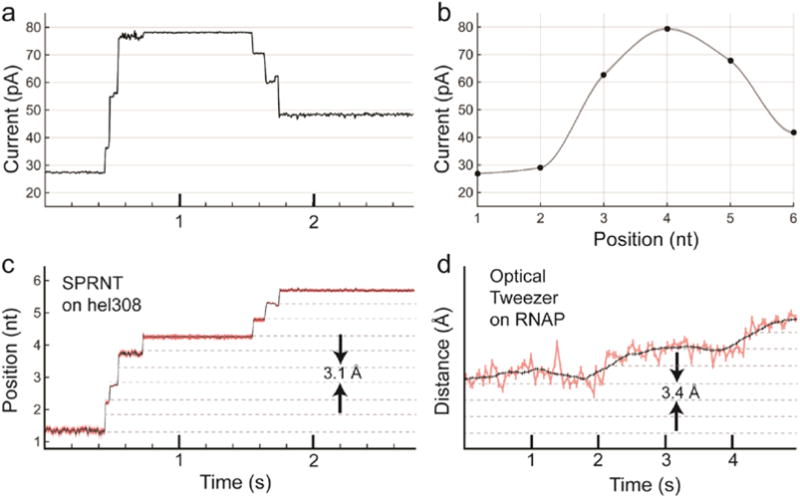
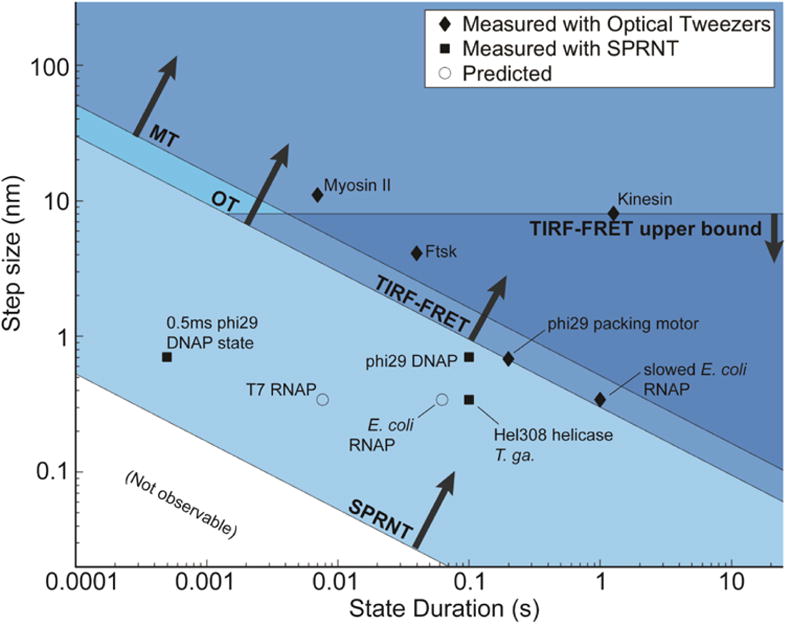
Nanopores are emerging as new single-molecule tools in the study of enzymes. Based on the progress in nanopore sequencing of DNA, a tool called Single-molecule Picometer Resolution Nanopore Tweezers (SPRNT) was developed to measure the movement of enzymes along DNA in real time. In this new method, an enzyme is loaded onto a DNA (or RNA) molecule. A single-stranded DNA end of this complex is drawn into a nanopore by an electrostatic potential that is applied across the pore. The single-stranded DNA passes through the pore's constriction until the enzyme comes into contact with the pore. Further progression of the DNA through the pore is then controlled by the enzyme. An ion current that flows through the pore's constriction is modulated by the DNA in the constriction. Analysis of ion current changes reveals the advance of the DNA with high spatiotemporal precision, thereby providing a real-time record of the enzyme's activity. Using an engineered version of the protein nanopore MspA, SPRNT has spatial resolution as small as 40pm at millisecond timescales, while simultaneously providing the DNA's sequence within the enzyme. In this chapter, SPRNT is introduced and its extraordinary potential is exemplified using the helicase Hel308. Two distinct substates are observed for each one-nucleotide advance; one of these about half-nucleotide long steps is ATP dependent and the other is ATP independent. The spatiotemporal resolution of this low-cost single-molecule technique lifts the study of enzymes to a new level of precision, enabling exploration of hitherto unobservable enzyme dynamics in real time.
Keywords: DNA sequencing; Enzyme kinetics; Force spectroscopy; High spatiotemporal resolution; In vitro; MspA; Single molecule; Single-nucleotide resolution.
© 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Adam G, Delbruck M. Reduction of dimensionality in biological diffusion processes. San Francisco: W.H: Freeman & Co; 1968.
-
- Adelman K, La Porta A, Santangelo TJ, Lis JT, Roberts JW, Wang MD. Single molecule analysis of RNA polymerase elongation reveals uniform kinetic behavior. Proceedings of the National Academy of Sciences of the United States of America. 2002;99(21):13538–13543. doi: 10.1073/pnas.212358999. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
