

Dual chemistry catalyzed by human acireductone dioxygenase
- PMID: 28062648
- PMCID: PMC5421613
- DOI: 10.1093/protein/gzw078
Dual chemistry catalyzed by human acireductone dioxygenase
Abstract
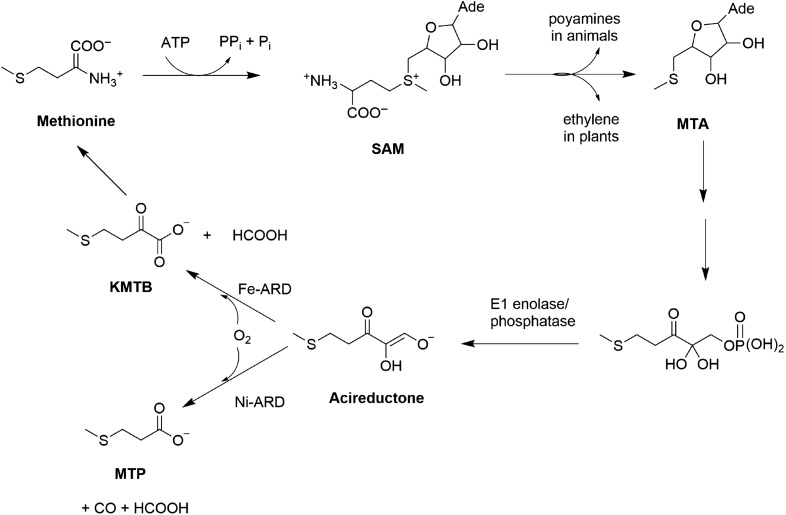
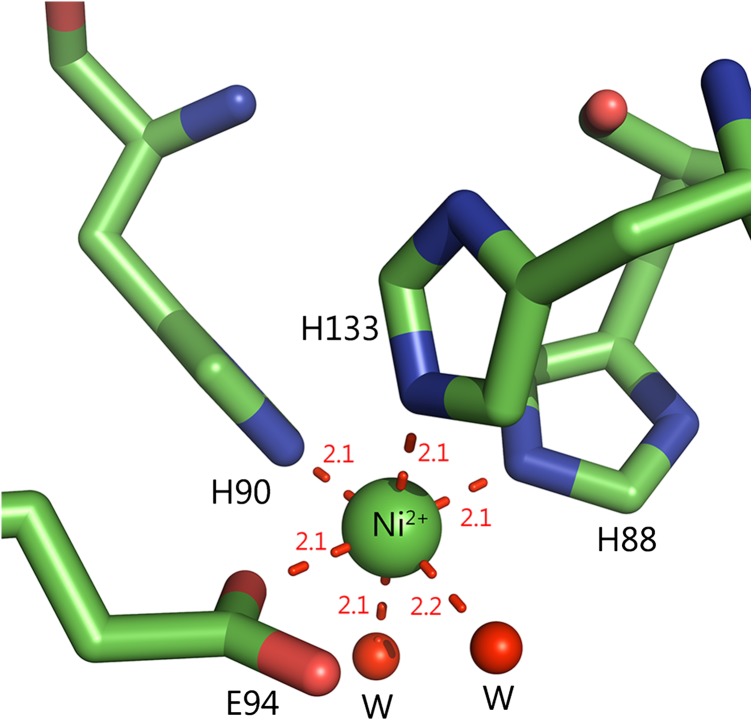
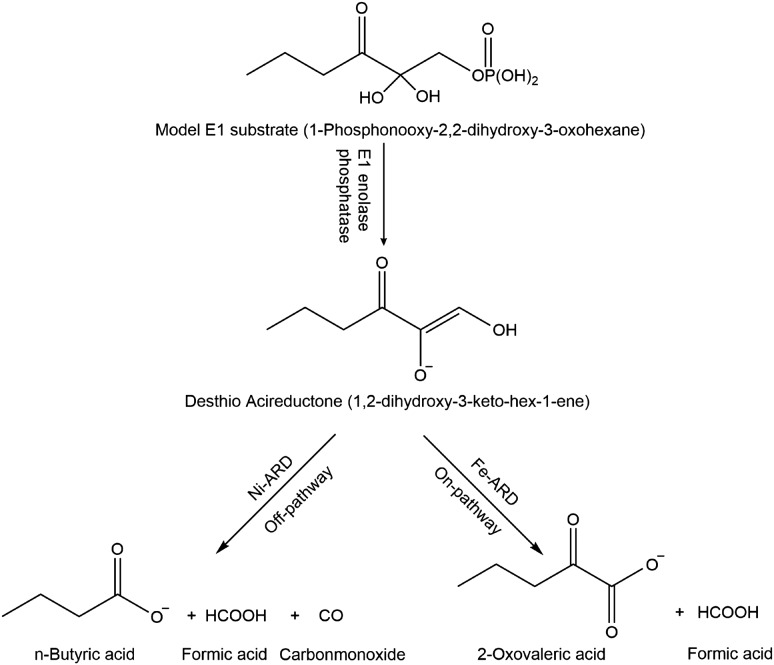
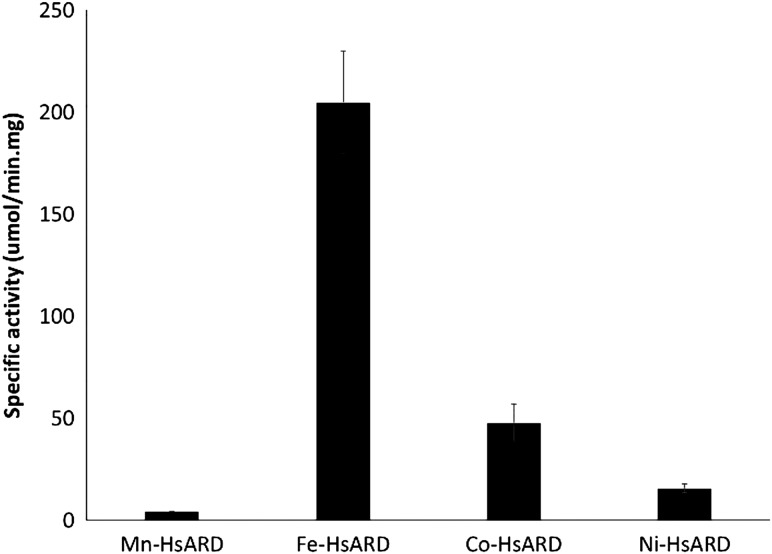
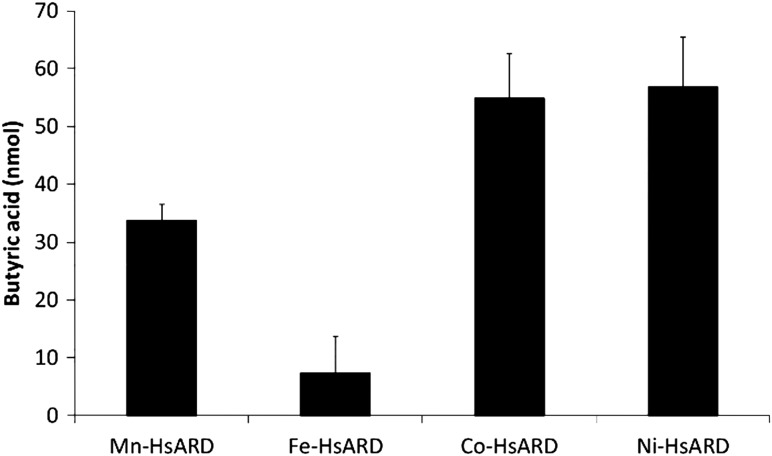
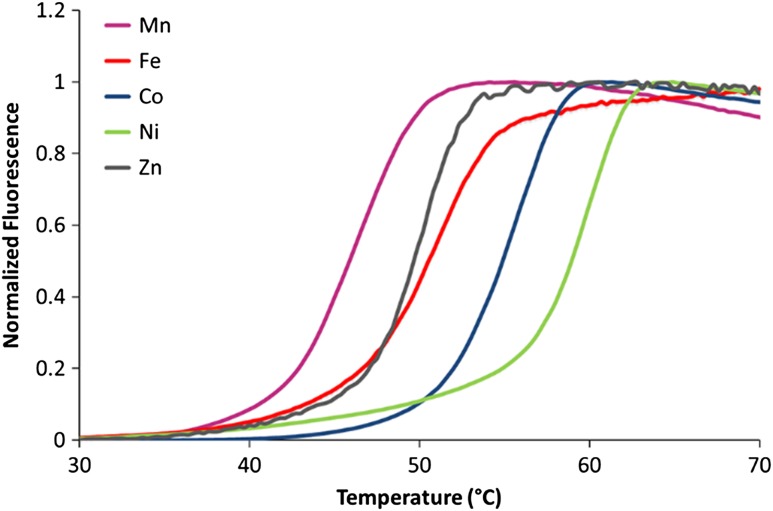
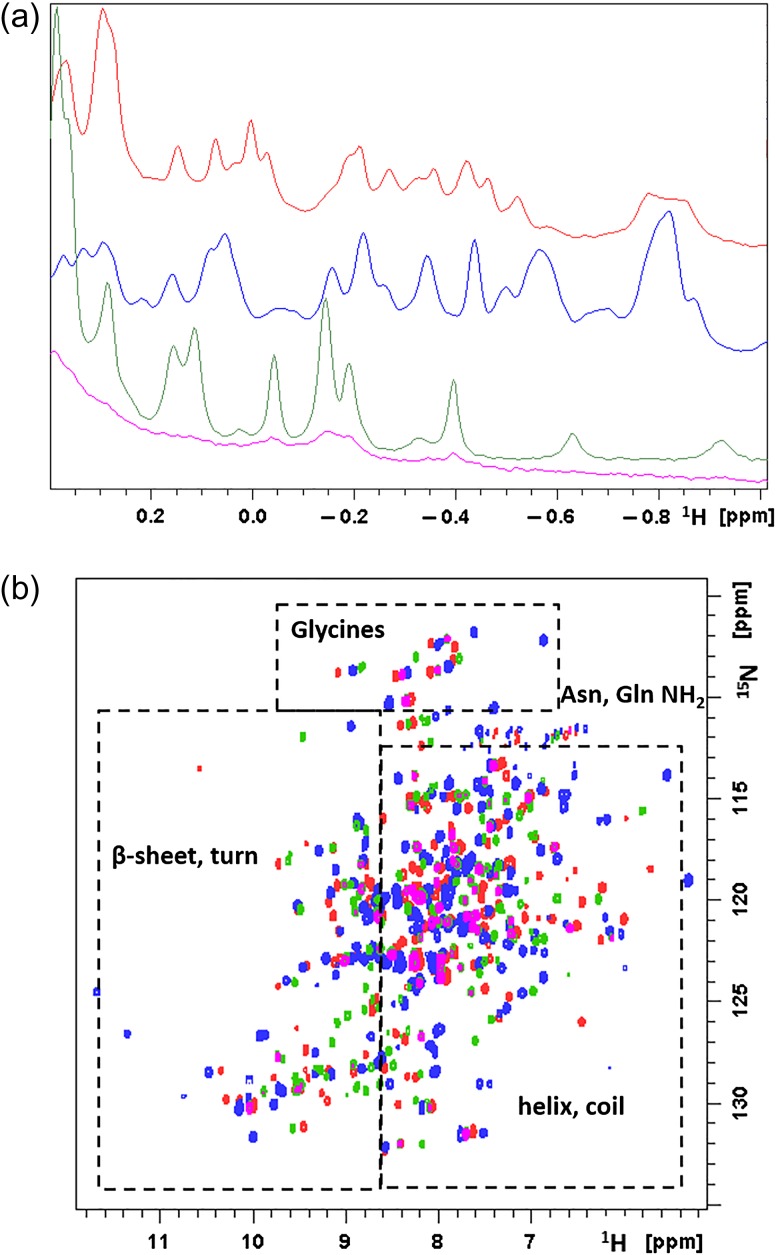
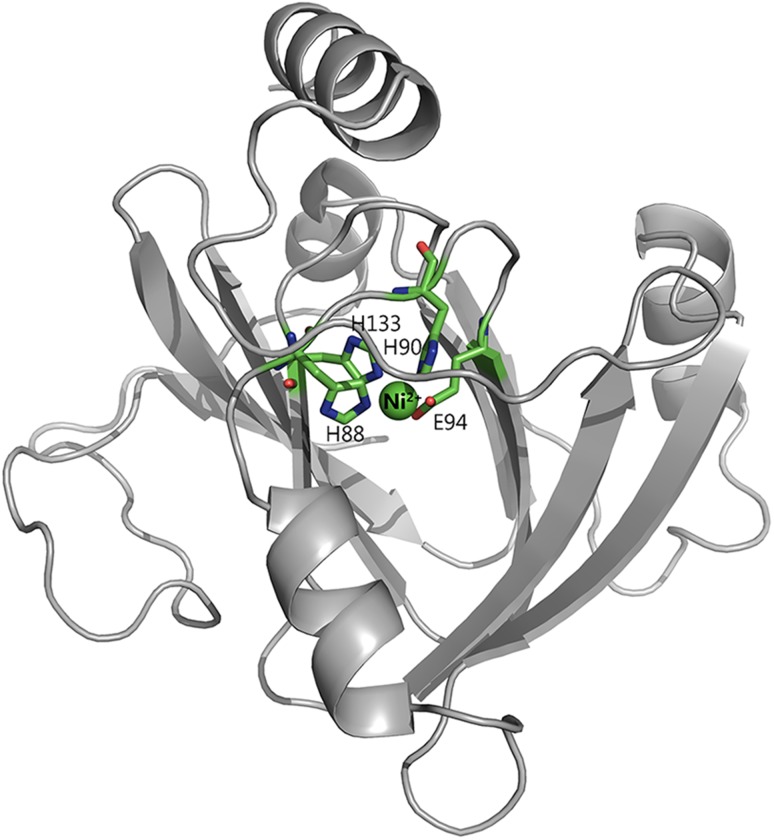
Acireductone dioxygenase (ARD) from the methionine salvage pathway of Klebsiella oxytoca is the only known naturally occurring metalloenzyme that catalyzes different reactions in vivo based solely on the identity of the divalent transition metal ion (Fe2+ or Ni2+) bound in the active site. The iron-containing isozyme catalyzes the cleavage of substrate 1,2-dihydroxy-3-keto-5-(thiomethyl)pent-1-ene (acireductone) by O2 to formate and the ketoacid precursor of methionine, whereas the nickel-containing isozyme uses the same substrates to catalyze an off-pathway shunt to form methylthiopropionate, carbon monoxide and formate. This dual chemistry was recently demonstrated in vitro by ARD from Mus musculus (MmARD), providing the first example of a mammalian ARD exhibiting metal-dependent catalysis. We now show that human ARD (HsARD) is also capable of metal-dependent dual chemistry. Recombinant HsARD was expressed and purified to obtain a homogeneous enzyme with a single transition metal ion bound. As with MmARD, the Fe2+-bound HsARD shows the highest activity and catalyzes on-pathway chemistry, whereas Ni2+, Co2+ or Mn2+ forms catalyze off-pathway chemistry. The thermal stability of the HsARD isozymes is a function of the metal ion identity, with Ni2+-bound HsARD being the most stable followed by Co2+ and Fe2+, and Mn2+-bound HsARD being the least stable. As with the bacterial ARD, solution NMR data suggest that HsARD isozymes can have significant structural differences depending upon the metal ion bound.
Keywords: Klebsiella oxytoca; acireductone; dual chemistry; mammalian; metal.
© The Author 2017. Published by Oxford University Press. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures

References
-
- Crans D.C., Yang L., Gaidamauskas E., Kahn R., Jin W. , Simonis U. (2003) In Paramagnetic Resonance of Metallobiomolecules (Telser, J., Ed.), American Chemical Society, Washington D.C.
-
- Dai Y., Pochapsky T.C. and Abeles R.H. (2001) Biochemistry, 40, 6379–6387. - PubMed
-
- Dai Y., Wensink P.C. and Abeles R.H. (1999) J. Biol. Chem., 274, 1193–1195. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
