

Translation of Expanded CGG Repeats into FMRpolyG Is Pathogenic and May Contribute to Fragile X Tremor Ataxia Syndrome
- PMID: 28065649
- PMCID: PMC5263258
- DOI: 10.1016/j.neuron.2016.12.016
Translation of Expanded CGG Repeats into FMRpolyG Is Pathogenic and May Contribute to Fragile X Tremor Ataxia Syndrome
Abstract
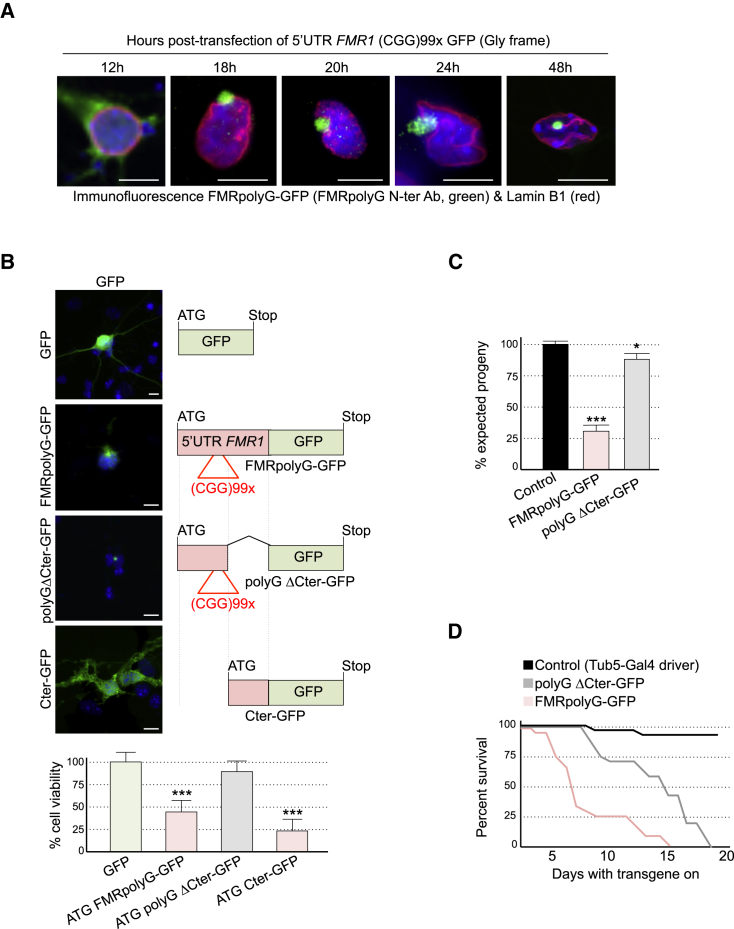
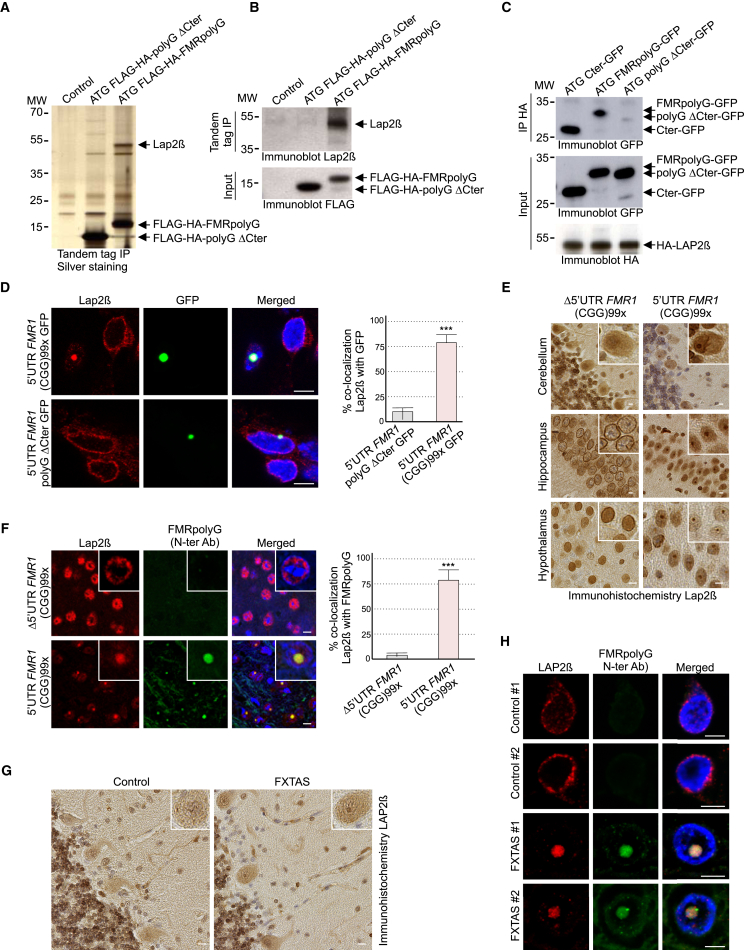
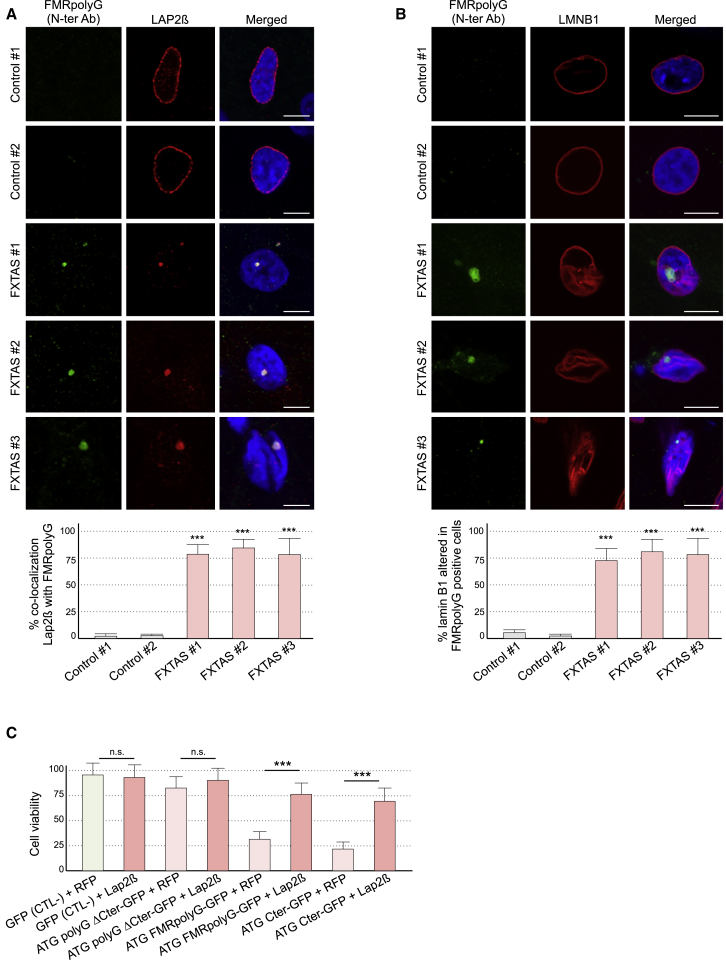
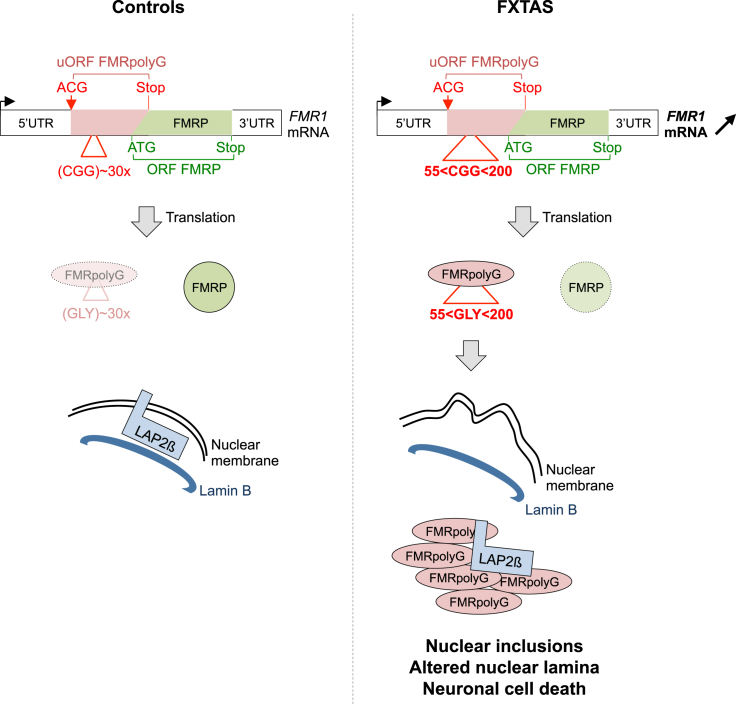
Fragile X-associated tremor/ataxia syndrome (FXTAS) is a neurodegenerative disorder caused by a limited expansion of CGG repeats in the 5' UTR of FMR1. Two mechanisms are proposed to cause FXTAS: RNA gain-of-function, where CGG RNA sequesters specific proteins, and translation of CGG repeats into a polyglycine-containing protein, FMRpolyG. Here we developed transgenic mice expressing CGG repeat RNA with or without FMRpolyG. Expression of FMRpolyG is pathogenic, while the sole expression of CGG RNA is not. FMRpolyG interacts with the nuclear lamina protein LAP2β and disorganizes the nuclear lamina architecture in neurons differentiated from FXTAS iPS cells. Finally, expression of LAP2β rescues neuronal death induced by FMRpolyG. Overall, these results suggest that translation of expanded CGG repeats into FMRpolyG alters nuclear lamina architecture and drives pathogenesis in FXTAS.
Keywords: RAN translation; microsatellite expansion; near-cognate codon; neurodegeneration.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
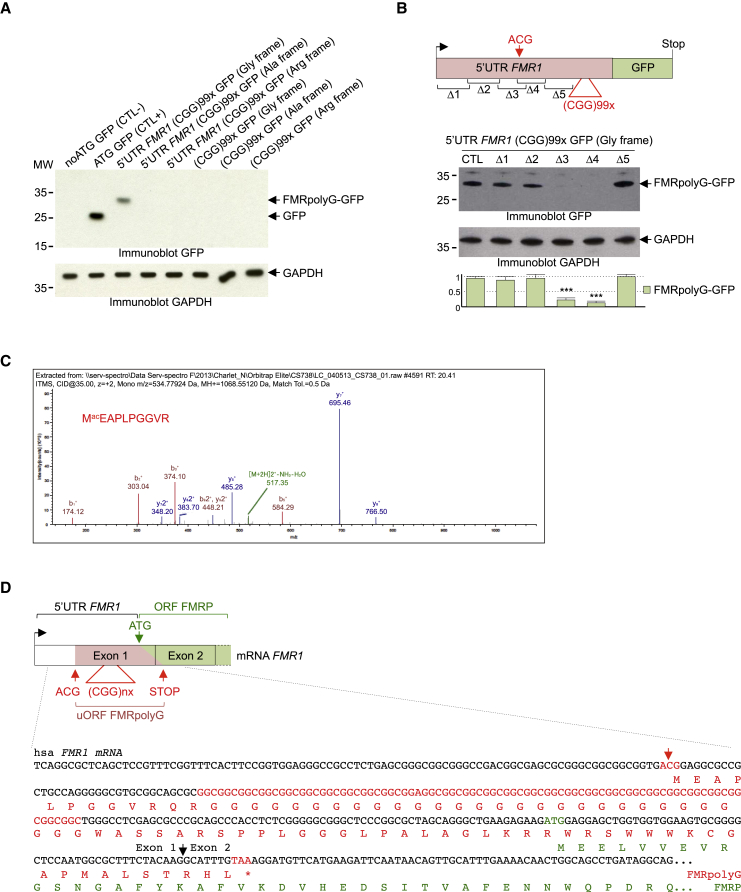
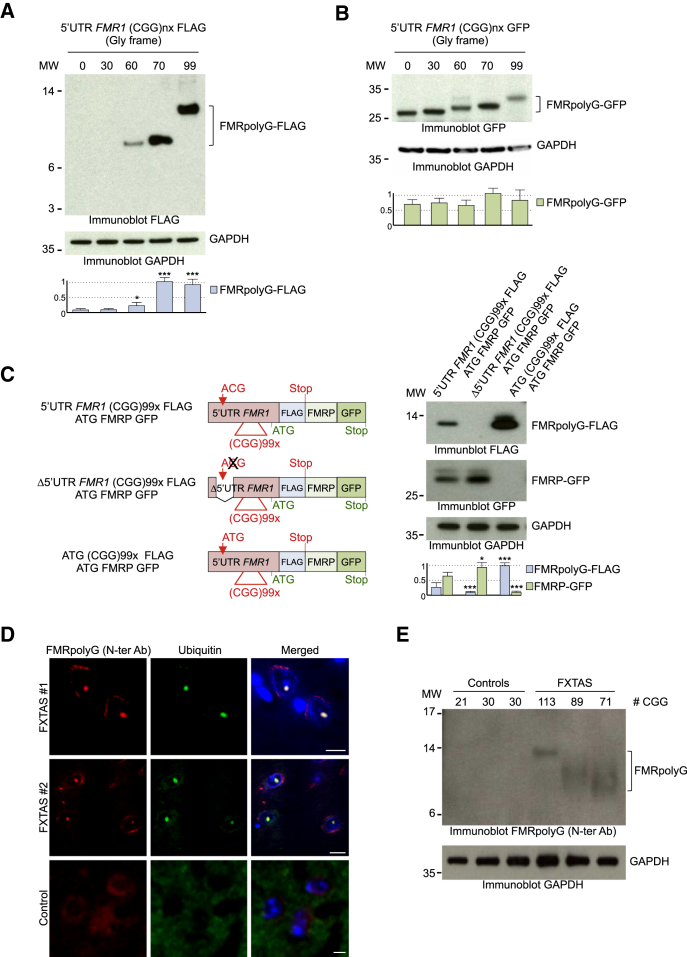
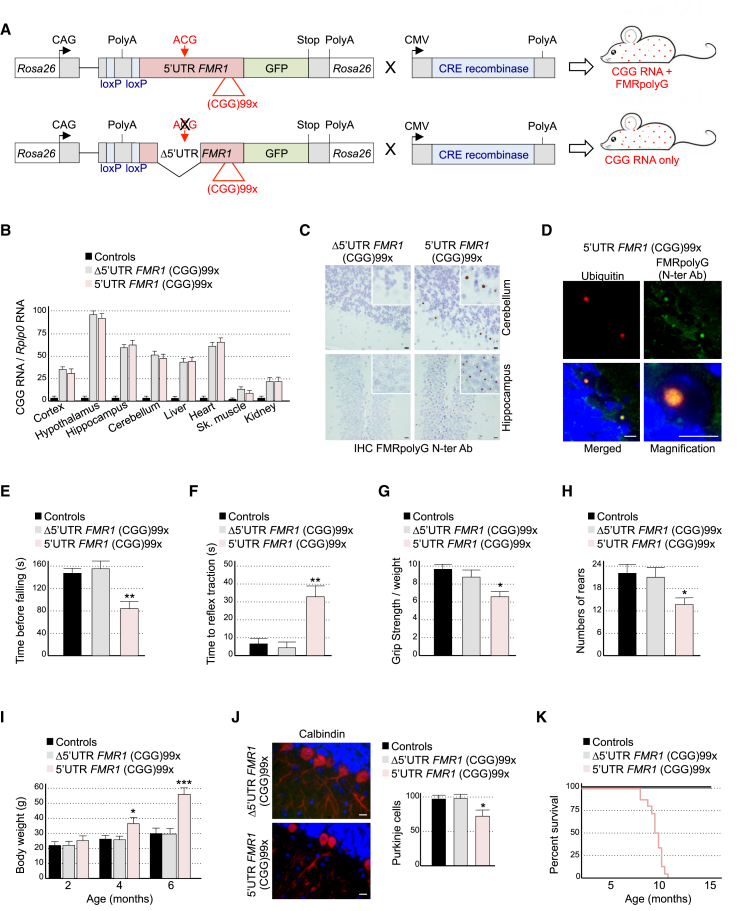
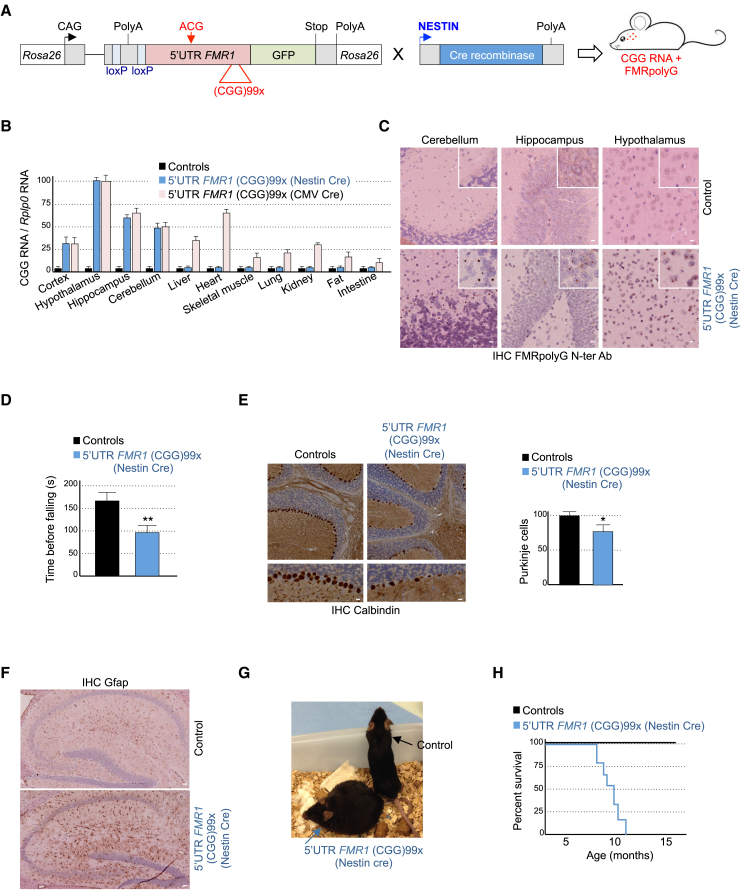
References
-
- Arocena D.G., Iwahashi C.K., Won N., Beilina A., Ludwig A.L., Tassone F., Schwartz P.H., Hagerman P.J. Induction of inclusion formation and disruption of lamin A/C structure by premutation CGG-repeat RNA in human cultured neural cells. Hum. Mol. Genet. 2005;14:3661–3671. - PubMed
-
- Ash P.E., Bieniek K.F., Gendron T.F., Caulfield T., Lin W.L., Dejesus-Hernandez M., van Blitterswijk M.M., Jansen-West K., Paul J.W., 3rd, Rademakers R. Unconventional translation of C9ORF72 GGGGCC expansion generates insoluble polypeptides specific to c9FTD/ALS. Neuron. 2013;77:639–646. - PMC - PubMed
-
- Buijsen R.A., Sellier C., Severijnen L.A., Oulad-Abdelghani M., Verhagen R.F., Berman R.F., Charlet-Berguerand N., Willemsen R., Hukema R.K. FMRpolyG-positive inclusions in CNS and non-CNS organs of a fragile X premutation carrier with fragile X-associated tremor/ataxia syndrome. Acta Neuropathol. Commun. 2014;2:162. - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
