

Network Dynamics Mediate Circadian Clock Plasticity
- PMID: 28065650
- PMCID: PMC5247339
- DOI: 10.1016/j.neuron.2016.12.022
Network Dynamics Mediate Circadian Clock Plasticity
Abstract
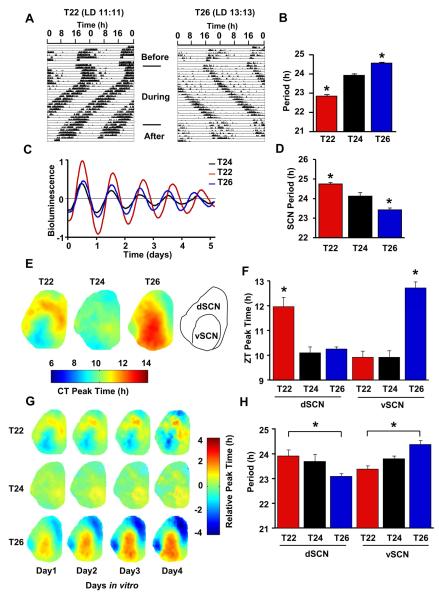
A circadian clock governs most aspects of mammalian behavior. Although its properties are in part genetically determined, altered light-dark environment can change circadian period length through a mechanism requiring de novo DNA methylation. We show here that this mechanism is mediated not via cell-autonomous clock properties, but rather through altered networking within the suprachiasmatic nuclei (SCN), the circadian "master clock," which is DNA methylated in region-specific manner. DNA methylation is necessary to temporally reorganize circadian phasing among SCN neurons, which in turn changes the period length of the network as a whole. Interruption of neural communication by inhibiting neuronal firing or by physical cutting suppresses both SCN reorganization and period changes. Mathematical modeling suggests, and experiments confirm, that this SCN reorganization depends upon GABAergic signaling. Our results therefore show that basic circadian clock properties are governed by dynamic interactions among SCN neurons, with neuroadaptations in network function driven by the environment.
Keywords: automation; autopatcher; in vivo; patch clamp; subcortical; thalamus; whole-cell.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures





References
-
- Albus H, Vansteensel MJ, Michel S, Block GD, Meijer JH. A GABAergic mechanism is necessary for coupling dissociable ventral and dorsal regional oscillators within the circadian clock. Current Biology. 2005;15:886–893. - PubMed
-
- Antle MC, Silver R. Orchestrating time: arrangements of the brain circadian clock. Trends in Neurosciences. 2005;28:145–151. - PubMed
-
- Aschoff J, Pohl H. Phase relations between a circadian rhythm and its zeitgeber within the range of entrainment. Naturwissenschaften. 1978;65:80–84. - PubMed
-
- Asher G, Schibler U. Crosstalk between components of circadian and metabolic cycles in mammals. Cell metabolism. 2011;13:125–137. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases