

PAMPs, PRRs, effectors and R-genes associated with citrus-pathogen interactions
- PMID: 28065920
- PMCID: PMC5571375
- DOI: 10.1093/aob/mcw238
PAMPs, PRRs, effectors and R-genes associated with citrus-pathogen interactions
Erratum in
-
PAMPs, PRRs, effectors and R-genes associated with citrus-pathogen interactions.Ann Bot. 2017 Oct 17;120(4):617. doi: 10.1093/aob/mcx076. Ann Bot. 2017. PMID: 28605397 Free PMC article. No abstract available.
Abstract
Background: Recent application of molecular-based technologies has considerably advanced our understanding of complex processes in plant-pathogen interactions and their key components such as PAMPs, PRRs, effectors and R-genes. To develop novel control strategies for disease prevention in citrus, it is essential to expand and consolidate our knowledge of the molecular interaction of citrus plants with their pathogens.
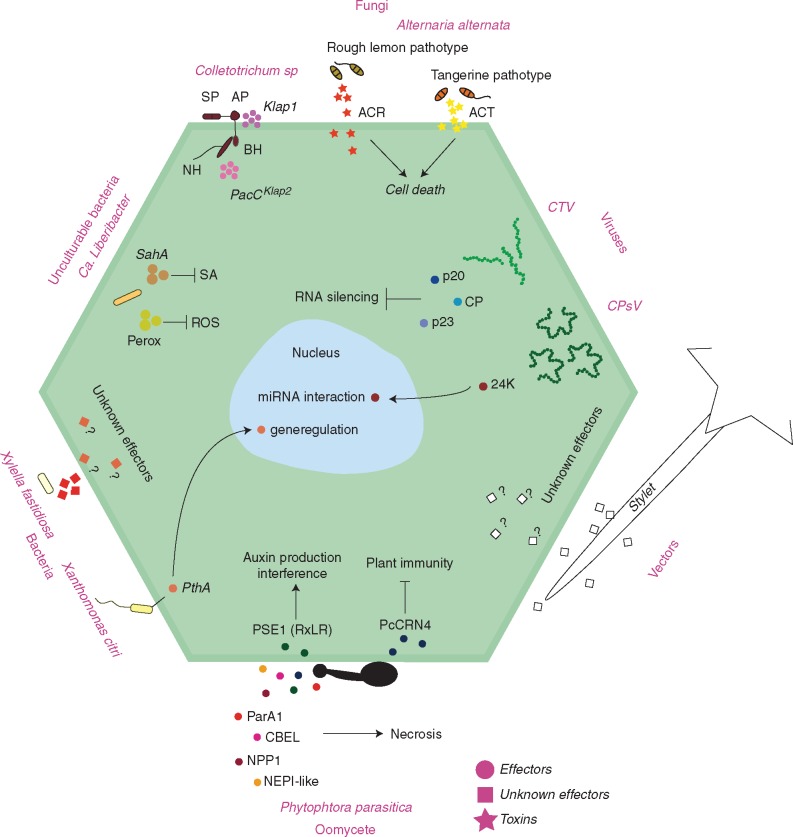
Scope: This review provides an overview of our understanding of citrus plant immunity, focusing on the molecular mechanisms involved in the interactions with viruses, bacteria, fungi, oomycetes and vectors related to the following diseases: tristeza, psorosis, citrus variegated chlorosis, citrus canker, huanglongbing, brown spot, post-bloom, anthracnose, gummosis and citrus root rot.
Keywords: Citrus immunity; and citrus root rot; anthracnose; brown spot; citrus canker; citrus psorosis; citrus variegated chlorosis (CVC); gummosis; huanglongbing (HLB); post-bloom; tristeza of citrus.
© The Author 2017. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For Permissions, please email: journals.permissions@oup.com
Figures
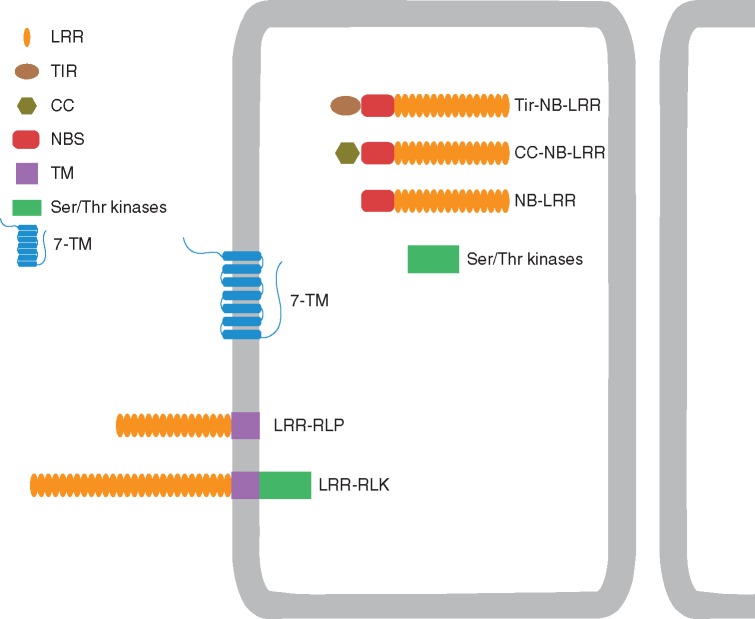
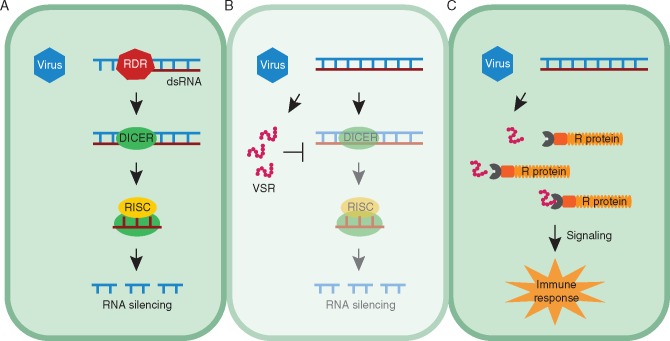
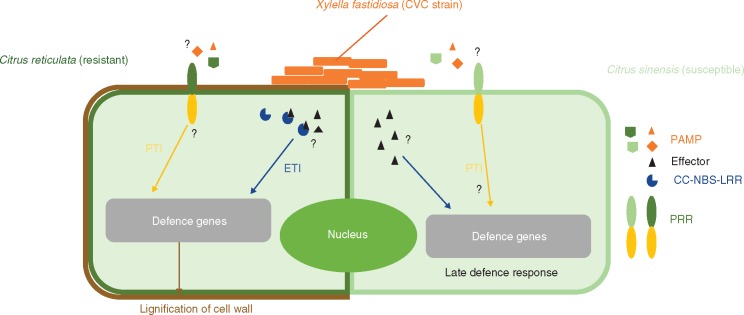
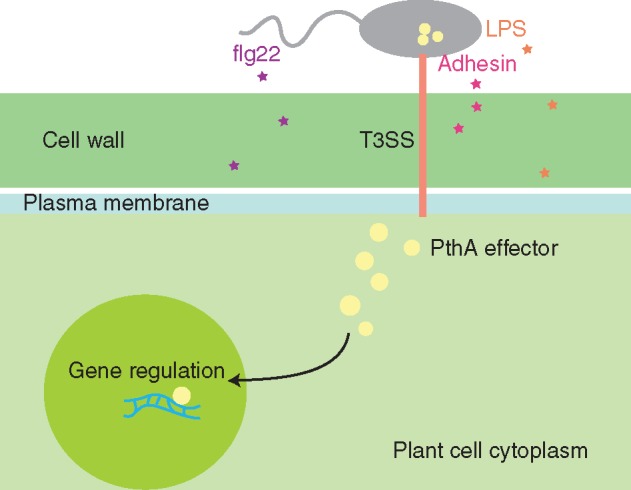



References
-
- Acevedo FE, Rivera-Vega LJ, Chung SH, Ray S, Felton GW.. 2015. Cues from chewing insects – the intersection of DAMPs, HAMPs, MAMPs and effectors. Current Opinion in Plant Biology 26: 80–86. - PubMed
-
- Adl SM, Simpson AGB, Farmer MA, et al. 2005. The new higher level classification of eukaryotes with emphasis on the taxonomy of protists. The Journal of Eukaryotic Microbiology 52: 399–451. - PubMed
-
- Akimitsu K, Peever TL, Timmer LW.. 2003. Molecular, ecological and evolutionary approaches to understanding Alternaria diseases of citrus. Molecular Plant Pathology 4: 435–446. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
