

CD36 deficiency impairs the small intestinal barrier and induces subclinical inflammation in mice
- PMID: 28066800
- PMCID: PMC5217470
- DOI: 10.1016/j.jcmgh.2016.09.001
CD36 deficiency impairs the small intestinal barrier and induces subclinical inflammation in mice
Abstract
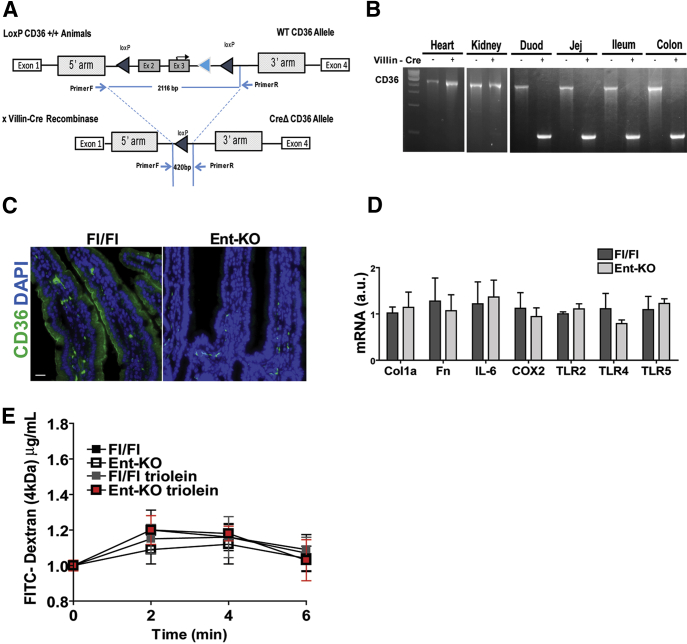
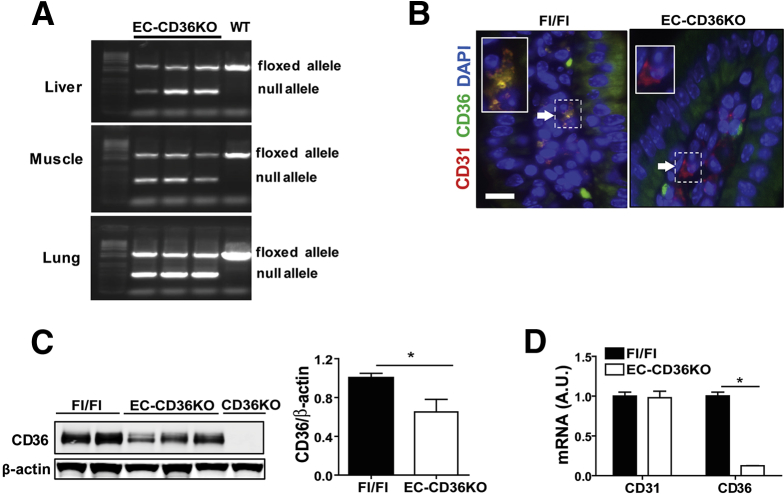
Background & aims: CD36 has immuno-metabolic actions and is abundant in the small intestine on epithelial, endothelial and immune cells. We examined the role of CD36 in gut homeostasis using mice null for CD36 (CD36KO) and with CD36 deletion specific to enterocytes (Ent-CD36KO) or endothelial cells (EC-CD36KO).
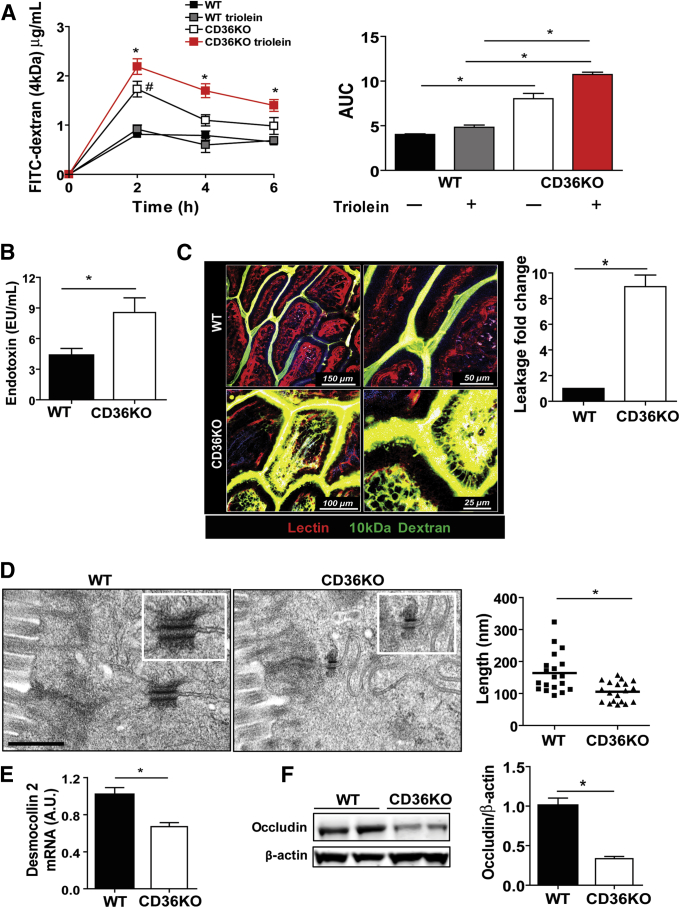
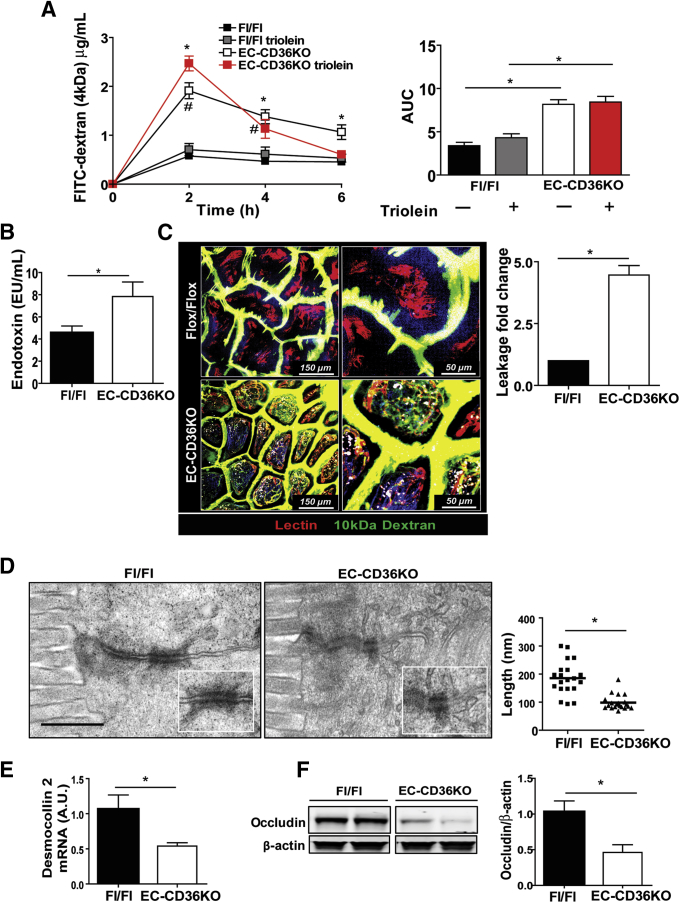
Methods: Intestinal morphology was evaluated using immunohistochemistry and electron microscopy (EM). Intestinal inflammation was determined from neutrophil infiltration and expression of cytokines, toll-like receptors and COX-2. Barrier integrity was assessed from circulating lipopolysaccharide (LPS) and dextran administered intragastrically. Epithelial permeability to luminal dextran was visualized using two photon microscopy.
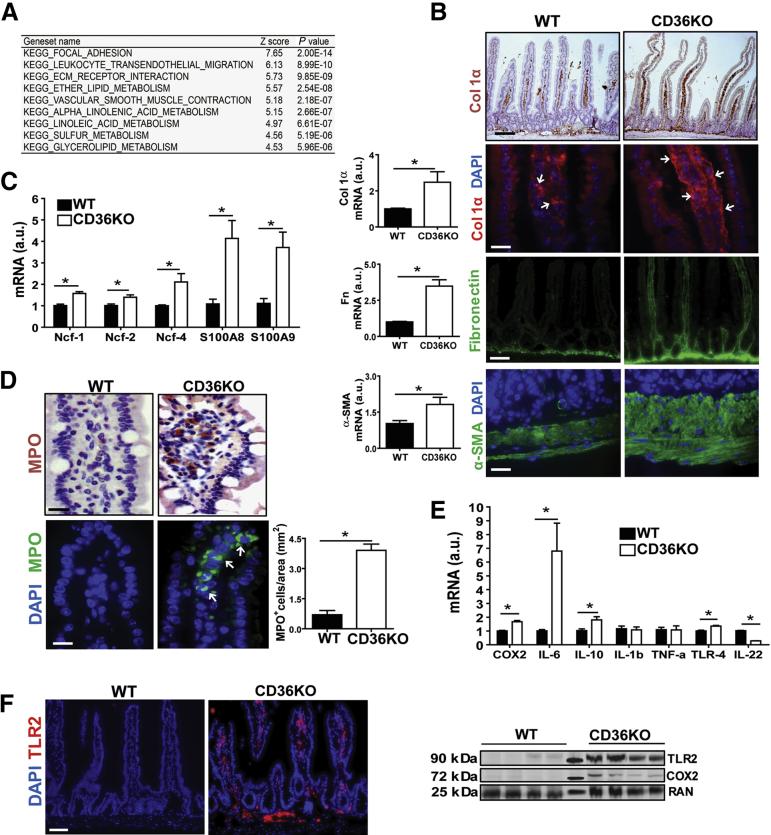
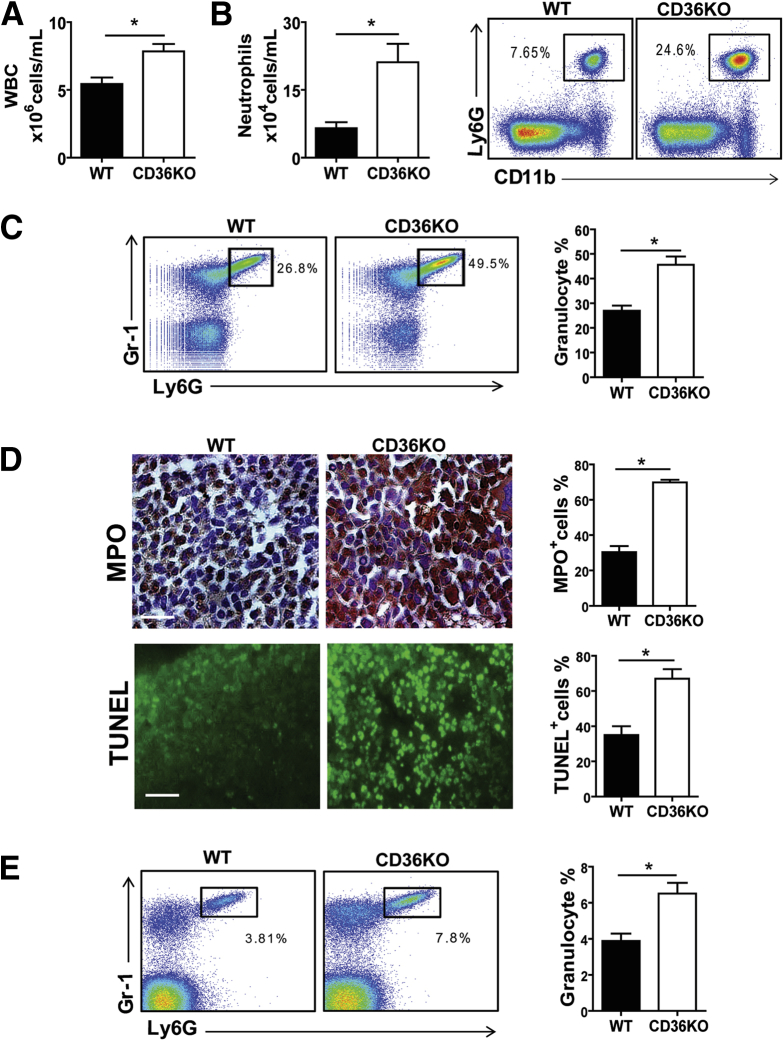
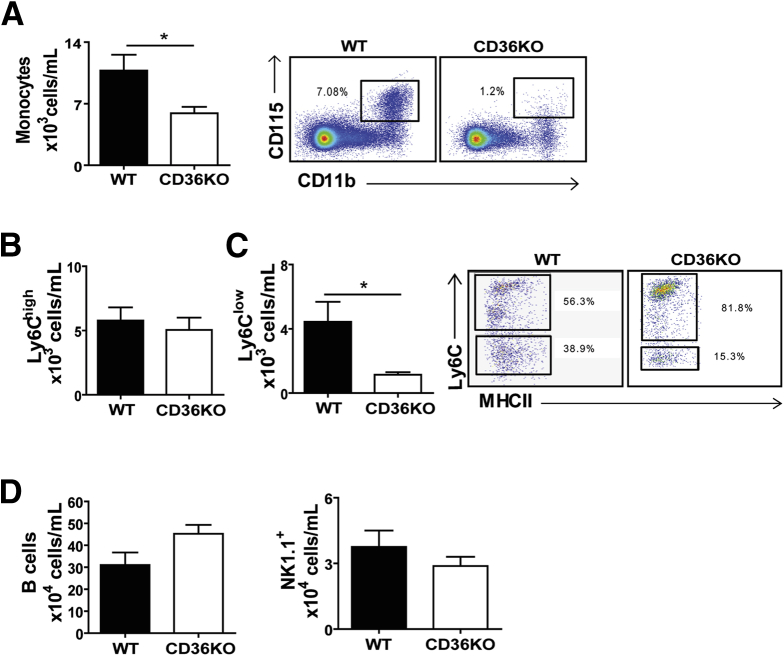
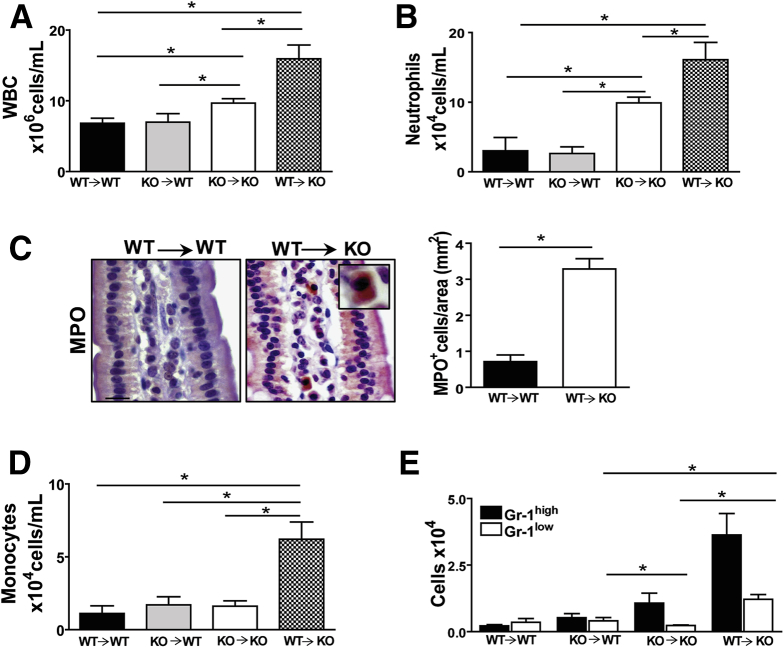
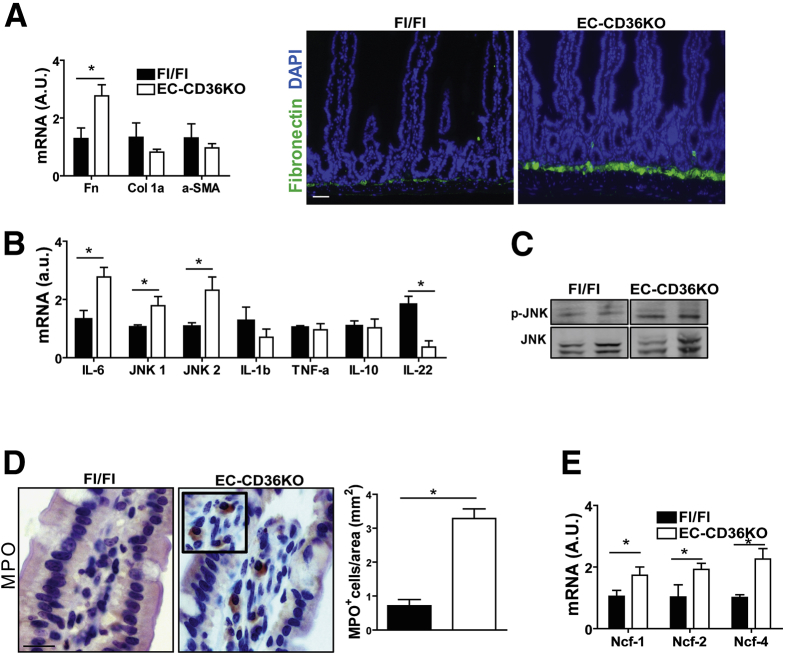
Results: The small intestines of CD36KO mice fed a chow diet showed several abnormalities including extracellular matrix (ECM) accumulation with increased expression of ECM proteins, evidence of neutrophil infiltration, inflammation and compromised barrier function. EM showed shortened desmosomes with decreased desmocollin 2 expression. Systemically, leukocytosis and neutrophilia were present together with 80% reduction of anti-inflammatory Ly6Clow monocytes. Bone marrow transplants supported the primary contribution of non-hematopoietic cells to the inflammatory phenotype. Specific deletion of endothelial but not of enterocyte CD36 reproduced many of the gut phenotypes of germline CD36KO mice including fibronectin deposition, increased interleukin 6, neutrophil infiltration, desmosome shortening and impaired epithelial barrier function.
Conclusions: CD36 loss results in chronic neutrophil infiltration of the gut, impairs barrier integrity and systemically causes subclinical inflammation. Endothelial cell CD36 deletion reproduces the major intestinal phenotypes. The findings suggest an important role of the endothelium in etiology of gut inflammation and loss of epithelial barrier integrity.
Keywords: Neutrophils; collagen; endothelium; fibronectin.
Conflict of interest statement
The authors have declared that no conflict of interest exists.
Figures
References
-
- Peterson L.W., Artis D. Intestinal epithelial cells: regulators of barrier function and immune homeostasis. Nat Rev Immunol. 2014;14:141–153. - PubMed
Grants and funding
- R01 DK060022/DK/NIDDK NIH HHS/United States
- R01 HL045095/HL/NHLBI NIH HHS/United States
- R21 AG046743/AG/NIA NIH HHS/United States
- P30 DK056341/DK/NIDDK NIH HHS/United States
- T32 HL007275/HL/NHLBI NIH HHS/United States
- DP1 DK109668/DK/NIDDK NIH HHS/United States
- R01 HL038180/HL/NHLBI NIH HHS/United States
- R01 AI049653/AI/NIAID NIH HHS/United States
- R37 HL045095/HL/NHLBI NIH HHS/United States
- R01 DK033301/DK/NIDDK NIH HHS/United States
- R37 HL038180/HL/NHLBI NIH HHS/United States
- R01 DK056260/DK/NIDDK NIH HHS/United States
- P30 DK052574/DK/NIDDK NIH HHS/United States
- R01 HL073029/HL/NHLBI NIH HHS/United States
- R01 DK095684/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous