

Why so narrow: Distribution of anti-sense regulated, type I toxin-antitoxin systems compared with type II and type III systems
- PMID: 28067598
- PMCID: PMC5367252
- DOI: 10.1080/15476286.2016.1272747
Why so narrow: Distribution of anti-sense regulated, type I toxin-antitoxin systems compared with type II and type III systems
Abstract
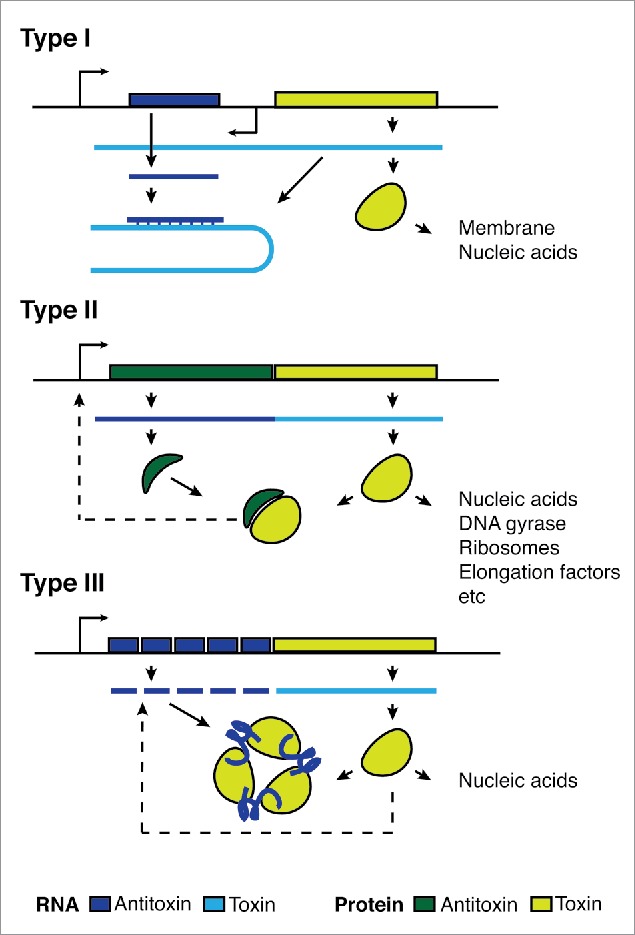
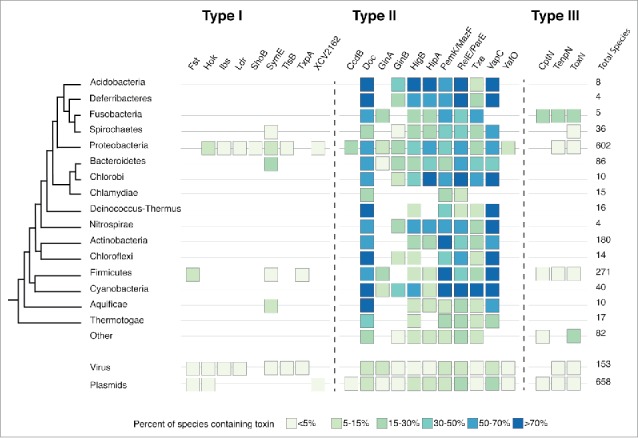
Toxin-antitoxin (TA) systems are gene modules that appear to be horizontally mobile across a wide range of prokaryotes. It has been proposed that type I TA systems, with an antisense RNA-antitoxin, are less mobile than other TAs that rely on direct toxin-antitoxin binding but no direct comparisons have been made. We searched for type I, II and III toxin families using iterative searches with profile hidden Markov models across phyla and replicons. The distribution of type I toxin families were comparatively narrow, but these patterns weakened with recently discovered families. We discuss how the function and phenotypes of TA systems as well as biases in our search methods may account for differences in their distribution.
Keywords: Antisense RNA; horizontal gene transfer; post-segregational killing; toxin-antitoxin systems.
Figures
References
-
- Van Melderen L, De Bast MS. Bacterial toxin-antitoxin systems: more than selfish entities? PLoS Genetics 2009; 5(3); PMID:19325885; http://dx.doi.org/15864262 10.1371/journal/pgen.1000437 - DOI - PMC - PubMed
-
- Gerdes K, Christensen SK, Løbner-Olesen A. Prokaryotic toxin-antitoxin stress response loci. Nat Rev Microbiol 2005; 3(5):371-82; PMID:15864262; http://dx.doi.org/ 10.1038/nrmicro1147 - DOI - PubMed
-
- Silvaggi JM, Perkins JB, Losick R. Small untranslated RNA antitoxin in Bacillus subtilis. J Bacteriol 2005; 187(19):6641-50; PMID:16166525; http://dx.doi.org/ 10.1128/JB.187.19.6641-6650.2005 - DOI - PMC - PubMed
-
- Kawano M, Reynolds AA, Miranda-Rios J, Storz G. Detection of 5′-and 3′-UTR-derived small RNAs and cis-encoded antisense RNAs in Escherichia coli. Nucleic Acids Res 2005; 33(3):1040-50; PMID:15718303; http://dx.doi.org/ 10.1093/nar/gki256 - DOI - PMC - PubMed
-
- Darfeuille F, Unoson C, Vogel Jr, Wagner EGH. An antisense RNA inhibits translation by competing with standby ribosomes. Mol Cell 2007; 26(3):381-92; PMID:17499044; http://dx.doi.org/ 10.1016/j.molcel.2007.04.003 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources