

Mechanisms to Avoid and Correct Erroneous Kinetochore-Microtubule Attachments
- PMID: 28067761
- PMCID: PMC5371994
- DOI: 10.3390/biology6010001
Mechanisms to Avoid and Correct Erroneous Kinetochore-Microtubule Attachments
Abstract
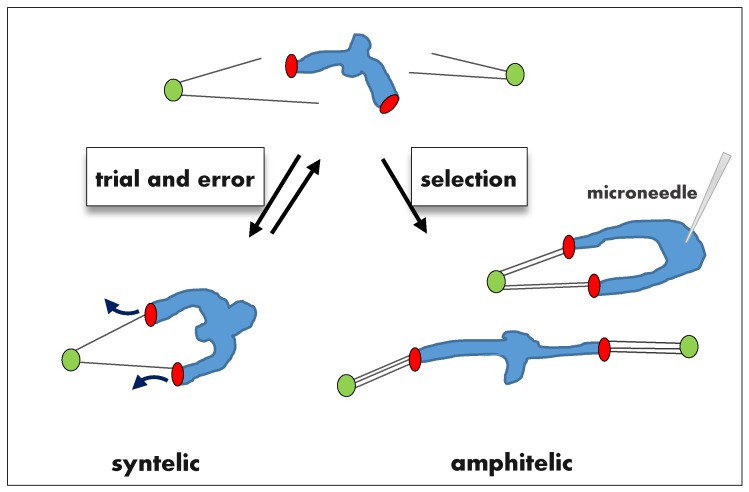
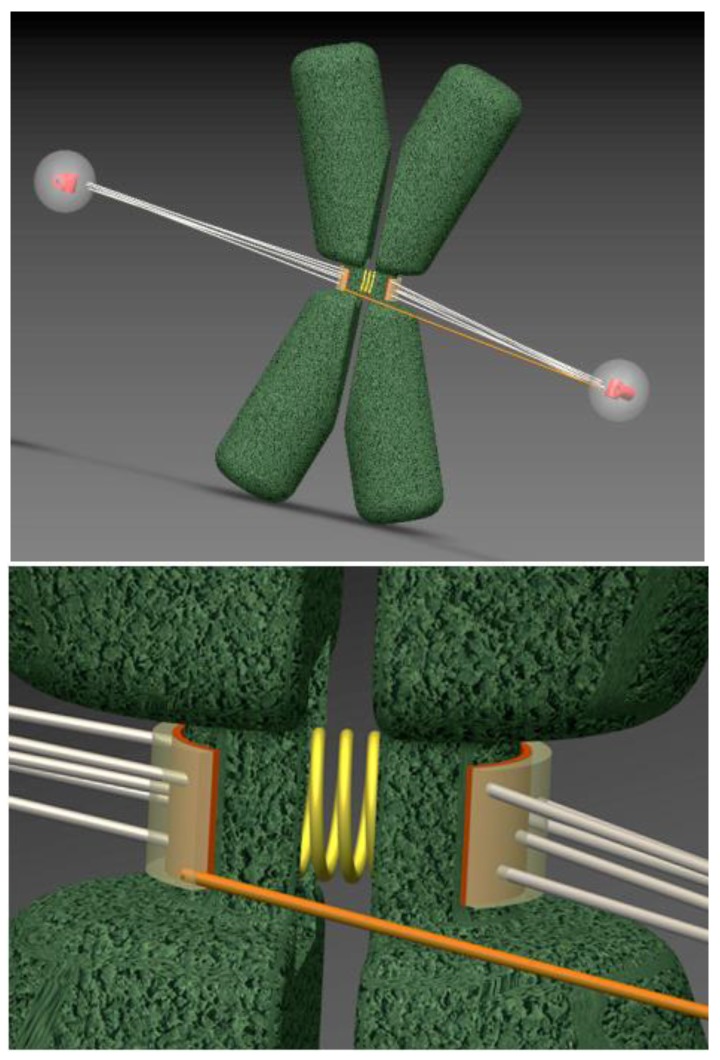
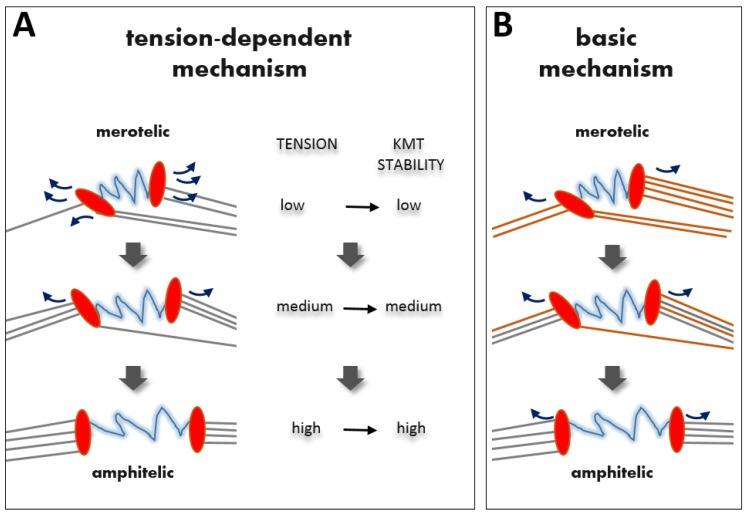
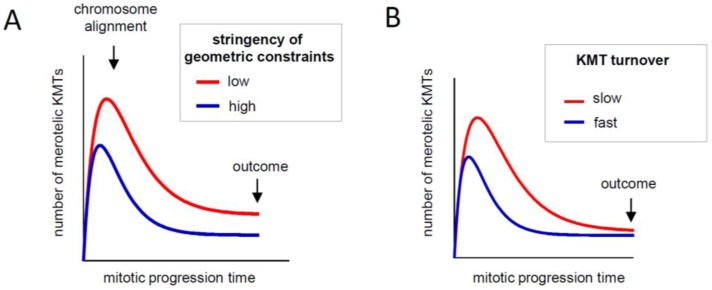
In dividing vertebrate cells multiple microtubules must connect to mitotic kinetochores in a highly stereotypical manner, with each sister kinetochore forming microtubule attachments to only one spindle pole. The exact sequence of events by which this goal is achieved varies considerably from cell to cell because of the variable locations of kinetochores and spindle poles, and randomness of initial microtubule attachments. These chance encounters with the kinetochores nonetheless ultimately lead to the desired outcome with high fidelity and in a limited time frame, providing one of the most startling examples of biological self-organization. This chapter discusses mechanisms that contribute to accurate chromosome segregation by helping dividing cells to avoid and resolve improper microtubule attachments.
Keywords: Aurora B kinase; kinetochore geometry; microtubule turnover; tension-dependent regulation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
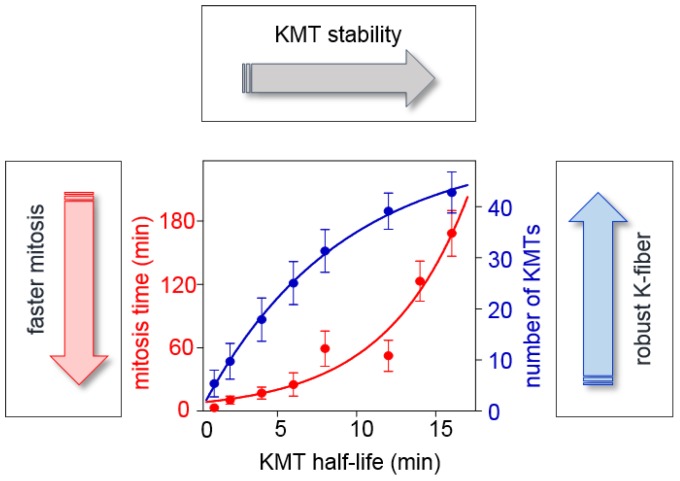
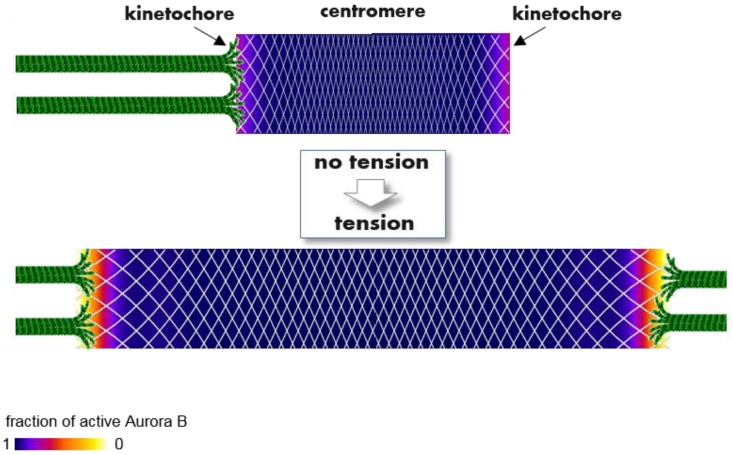
References
-
- Nicholson J.M., Cimini D. Link between aneuploidy and chromosome instability. Int. Rev. Cell Mol. Biol. 2015;315:299–317. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
