

Prophage-mediated defence against viral attack and viral counter-defence
- PMID: 28067906
- PMCID: PMC5508108
- DOI: 10.1038/nmicrobiol.2016.251
Prophage-mediated defence against viral attack and viral counter-defence
Abstract
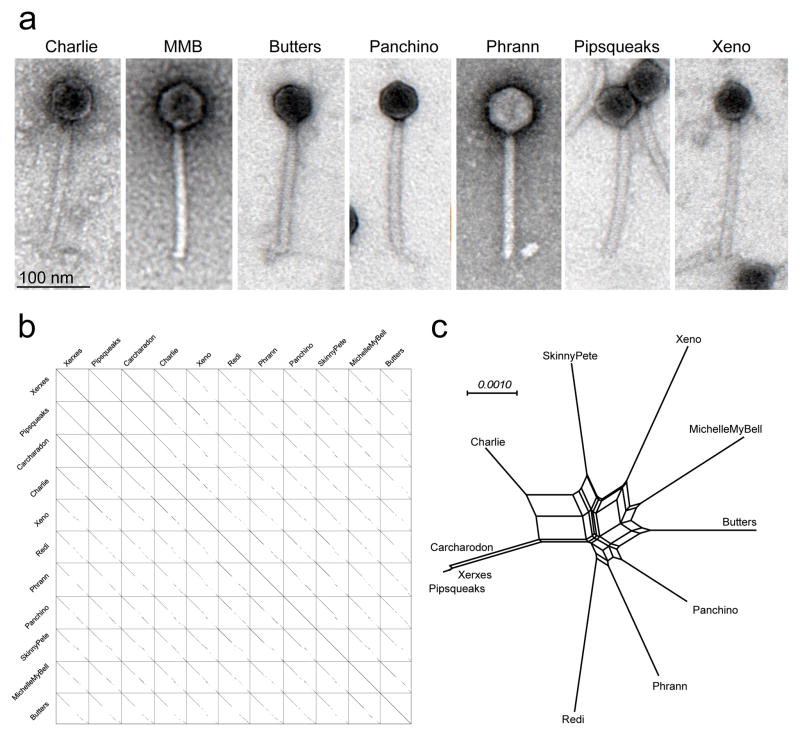
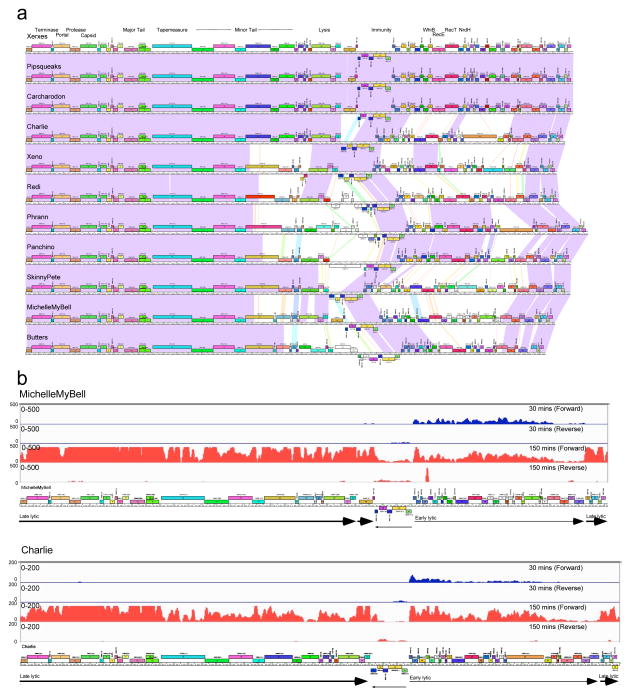
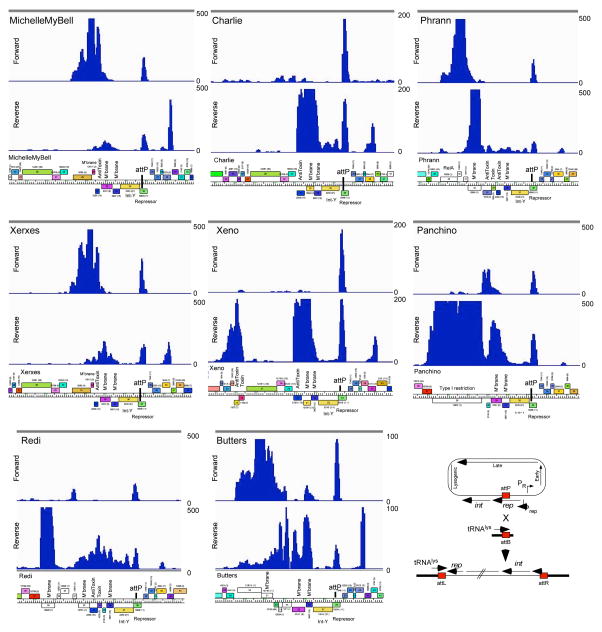
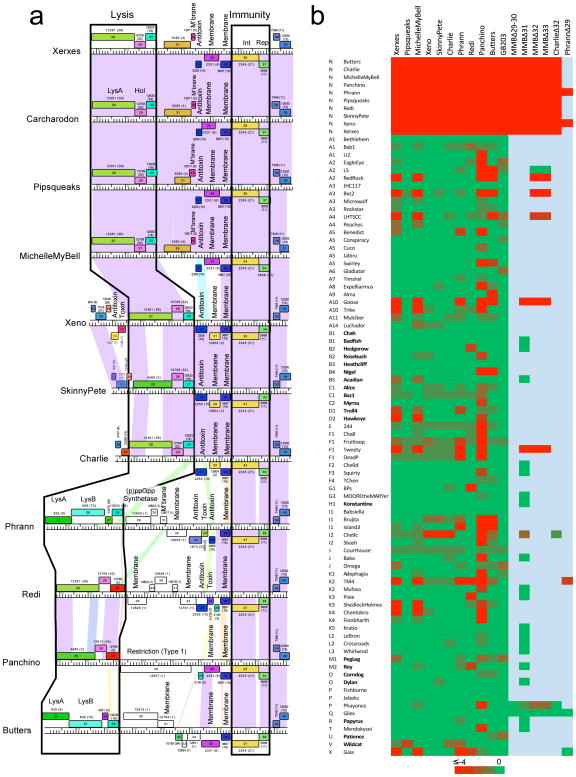
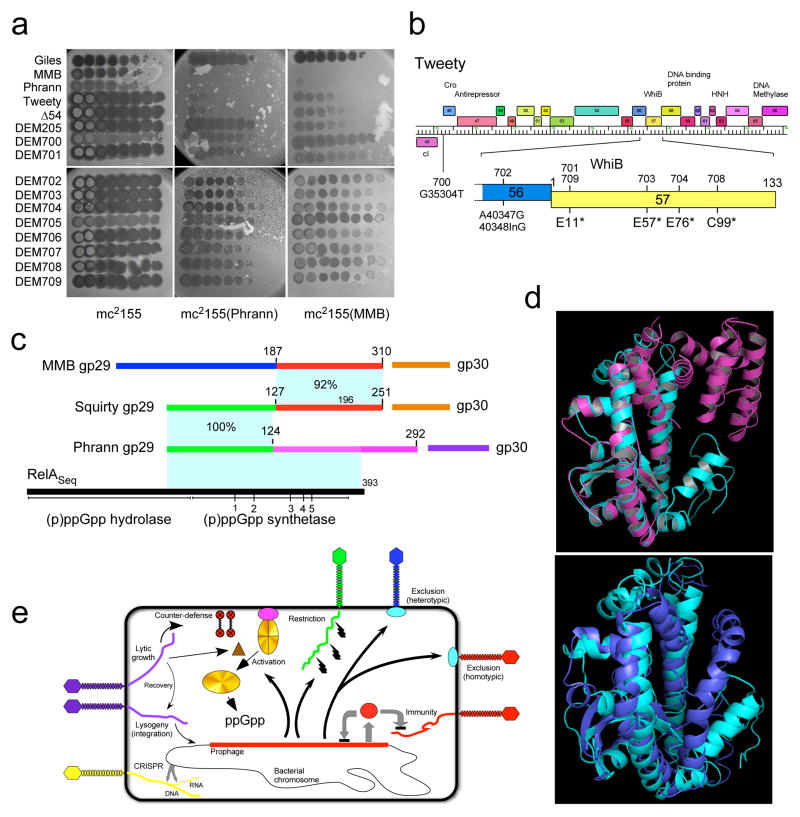
Temperate phages are common, and prophages are abundant residents of sequenced bacterial genomes. Mycobacteriophages are viruses that infect mycobacterial hosts including Mycobacterium tuberculosis and Mycobacterium smegmatis, encompass substantial genetic diversity and are commonly temperate. Characterization of ten Cluster N temperate mycobacteriophages revealed at least five distinct prophage-expressed viral defence systems that interfere with the infection of lytic and temperate phages that are either closely related (homotypic defence) or unrelated (heterotypic defence) to the prophage. Target specificity is unpredictable, ranging from a single target phage to one-third of those tested. The defence systems include a single-subunit restriction system, a heterotypic exclusion system and a predicted (p)ppGpp synthetase, which blocks lytic phage growth, promotes bacterial survival and enables efficient lysogeny. The predicted (p)ppGpp synthetase coded by the Phrann prophage defends against phage Tweety infection, but Tweety codes for a tetrapeptide repeat protein, gp54, which acts as a highly effective counter-defence system. Prophage-mediated viral defence offers an efficient mechanism for bacterial success in host-virus dynamics, and counter-defence promotes phage co-evolution.
Figures

References
-
- Hambly E, Suttle CA. The viriosphere, diversity, and genetic exchange within phage communities. Curr Opin Microbiol. 2005;8:444–450. - PubMed
-
- Abedon ST. Phage evolution and ecology. Adv Appl Microbiol. 2009;67:1–45. - PubMed
-
- Hendrix RW. Bacteriophages: evolution of the majority. Theor Popul Biol. 2002;61:471–480. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
