

Derepression of the DNA Methylation Machinery of the Gata1 Gene Triggers the Differentiation Cue for Erythropoiesis
- PMID: 28069743
- PMCID: PMC5376630
- DOI: 10.1128/MCB.00592-16
Derepression of the DNA Methylation Machinery of the Gata1 Gene Triggers the Differentiation Cue for Erythropoiesis
Abstract
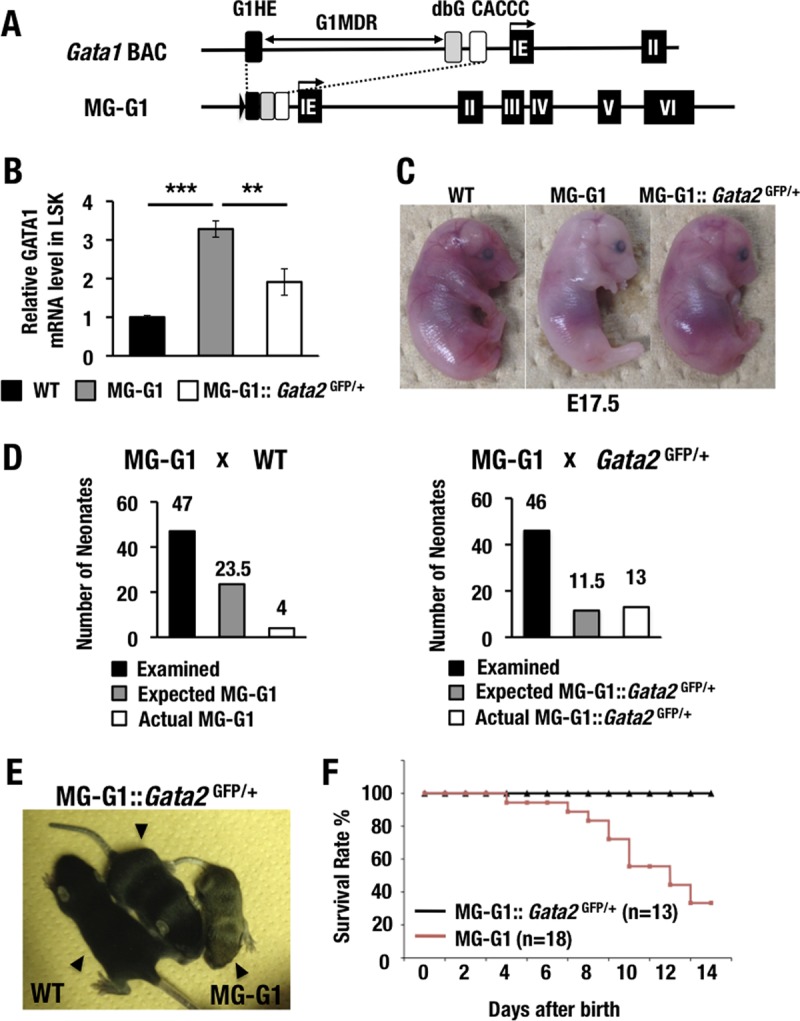
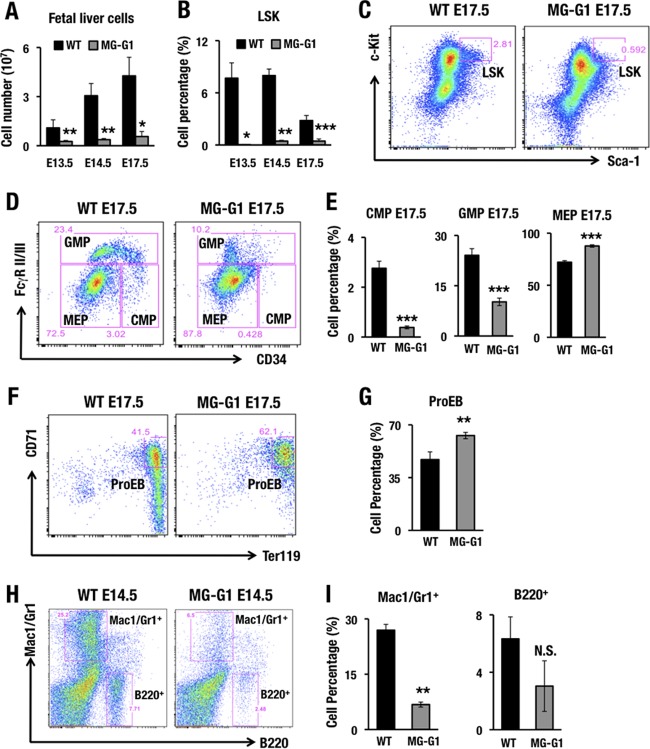
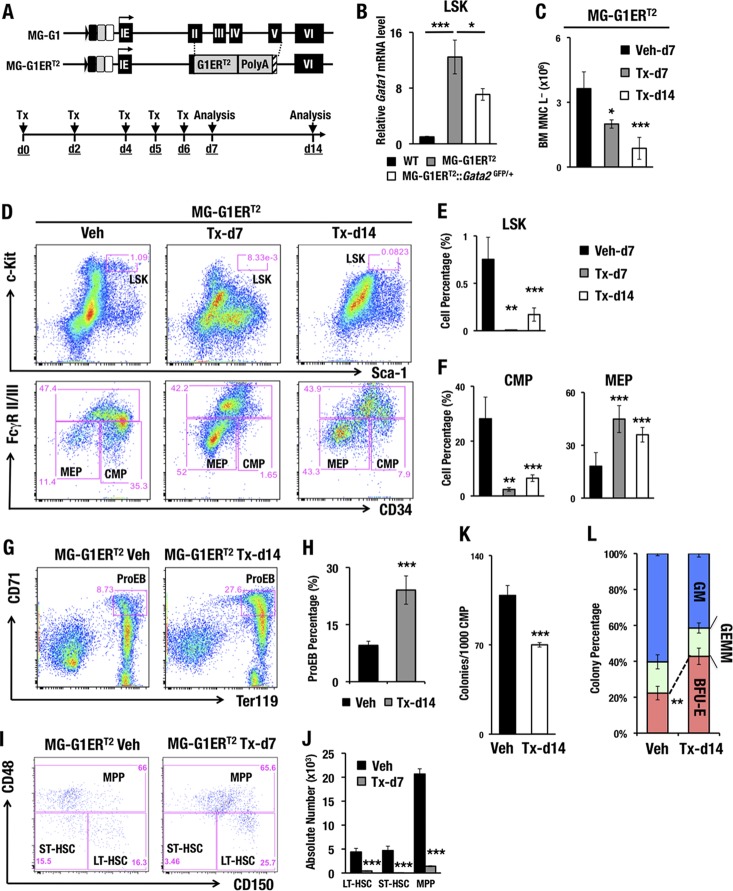
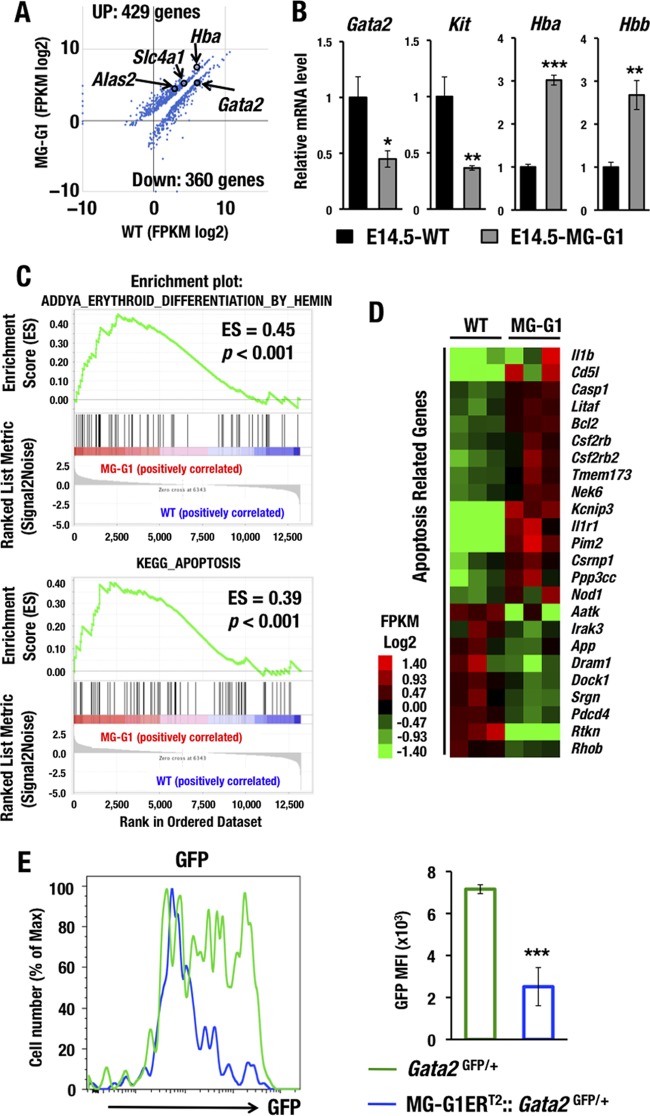
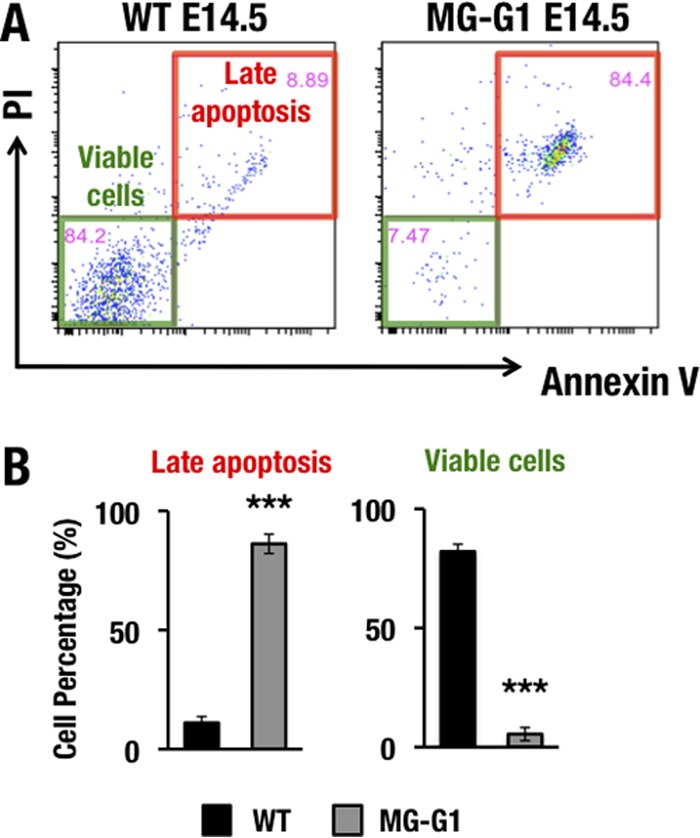
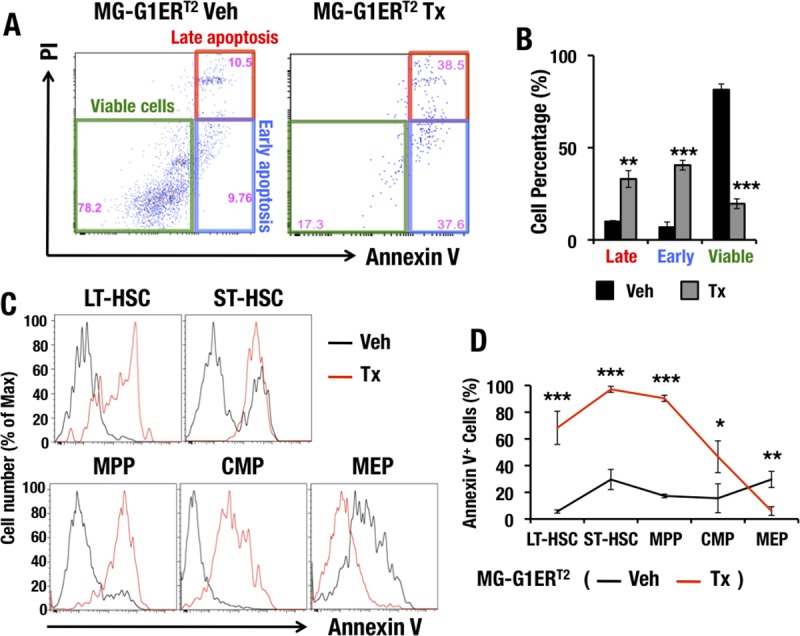
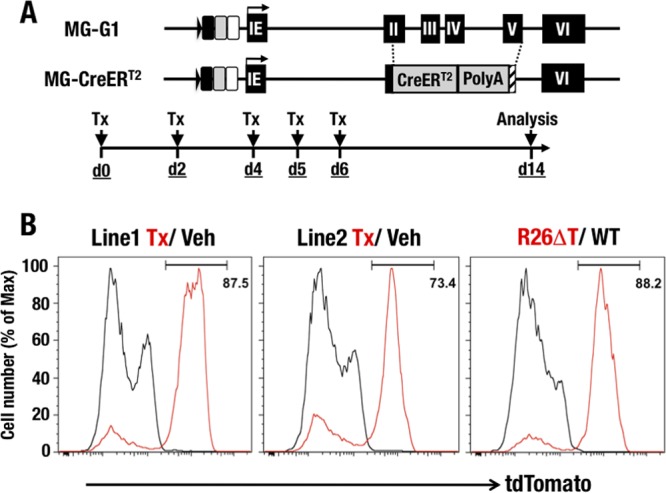
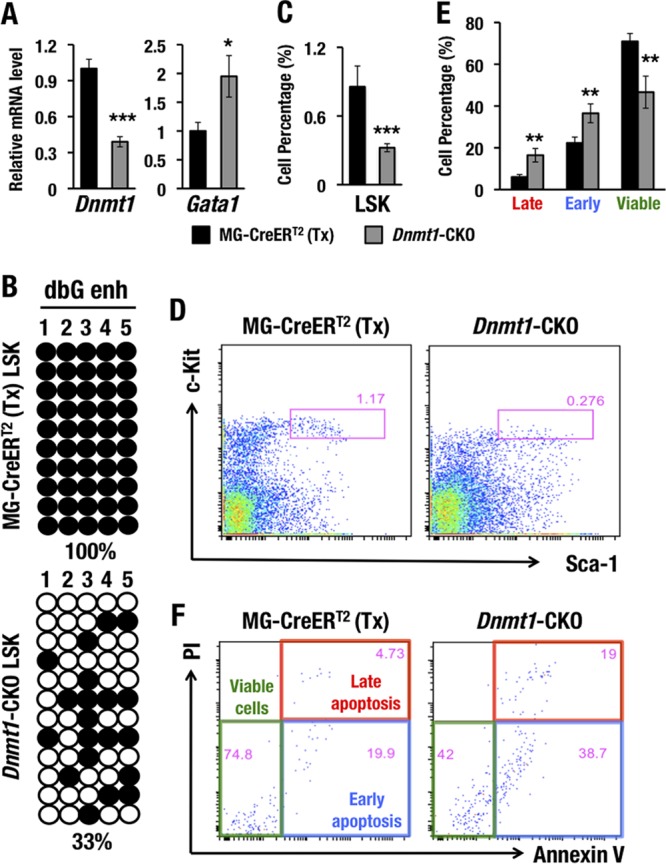
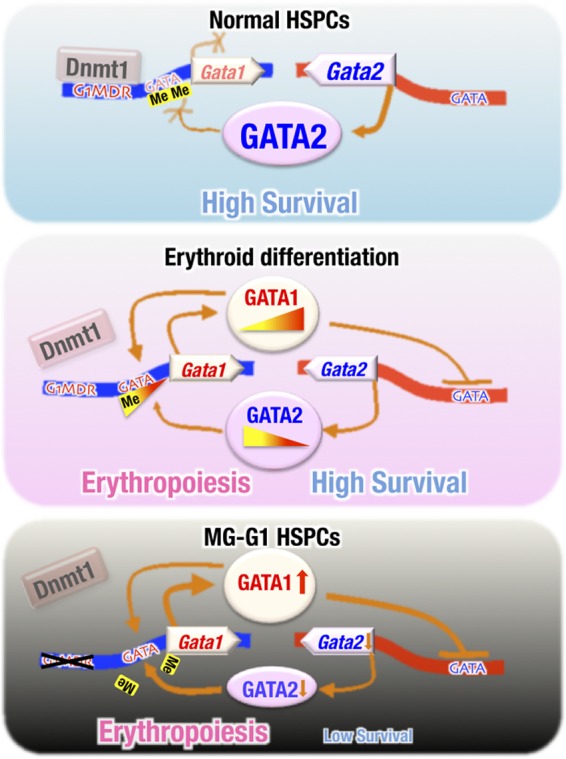
GATA1 is a critical regulator of erythropoiesis. While the mechanisms underlying the high-level expression of GATA1 in maturing erythroid cells have been studied extensively, the initial activation of the Gata1 gene in early hematopoietic progenitors remains to be elucidated. We previously identified a hematopoietic stem and progenitor cell (HSPC)-specific silencer element (the Gata1 methylation-determining region [G1MDR]) that recruits DNA methyltransferase 1 (Dnmt1) and provokes methylation of the Gata1 gene enhancer. In the present study, we hypothesized that removal of the G1MDR-mediated silencing machinery is the molecular basis of the initial activation of the Gata1 gene and erythropoiesis. To address this hypothesis, we generated transgenic mouse lines harboring a Gata1 bacterial artificial chromosome in which the G1MDR was deleted. The mice exhibited abundant GATA1 expression in HSPCs, in a GATA2-dependent manner. The ectopic GATA1 expression repressed Gata2 transcription and induced erythropoiesis and apoptosis of HSPCs. Furthermore, genetic deletion of Dnmt1 in HSPCs activated Gata1 expression and depleted HSPCs, thus recapitulating the HSC phenotype associated with GATA1 gain of function. These results demonstrate that the G1MDR holds the key to HSPC maintenance and suggest that release from this suppressive mechanism is a fundamental requirement for subsequent initiation of erythroid differentiation.
Keywords: Gata1 gene regulation; erythropoiesis; hematopoietic stem and progenitor cell.
Copyright © 2017 American Society for Microbiology.
Figures
References
-
- Takahashi S, Komeno T, Suwabe N, Yoh K, Nakajima O, Nishimura S, Kuroha T, Nagasawa T, Yamamoto M. 1998. Role of GATA-1 in proliferation and differentiation of definitive erythroid and megakaryocytic cells in vivo. Blood 92:434–442. - PubMed
-
- Zon LI, Yamaguchi Y, Yee K, Albee EA, Kimura A, Bennett JC, Orkin SH, Ackerman SJ. 1993. Expression of mRNA for the GATA-binding proteins in human eosinophils and basophils: potential role in gene transcription. Blood 81:3234–3241. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases