

Interaction of the CD43 Sialomucin with the Mycobacterium tuberculosis Cpn60.2 Chaperonin Leads to Tumor Necrosis Factor Alpha Production
- PMID: 28069816
- PMCID: PMC5328480
- DOI: 10.1128/IAI.00915-16
Interaction of the CD43 Sialomucin with the Mycobacterium tuberculosis Cpn60.2 Chaperonin Leads to Tumor Necrosis Factor Alpha Production
Abstract
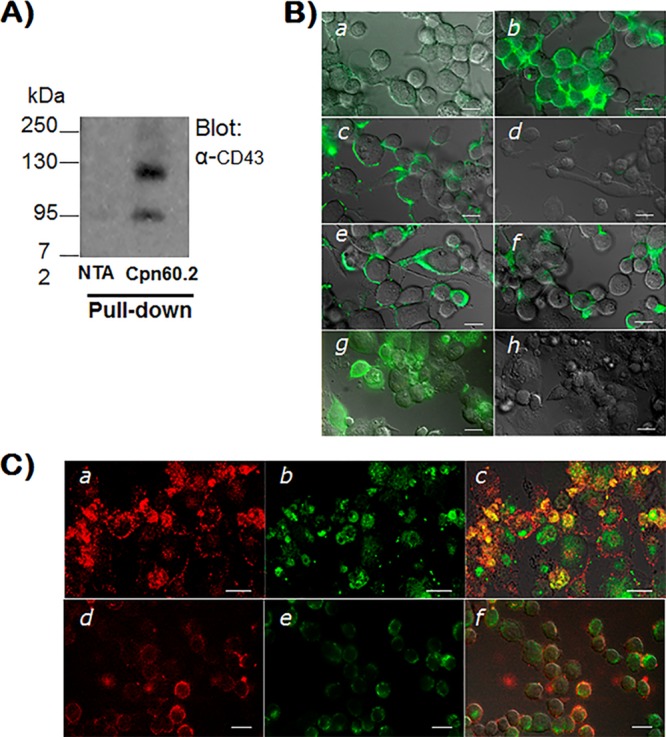
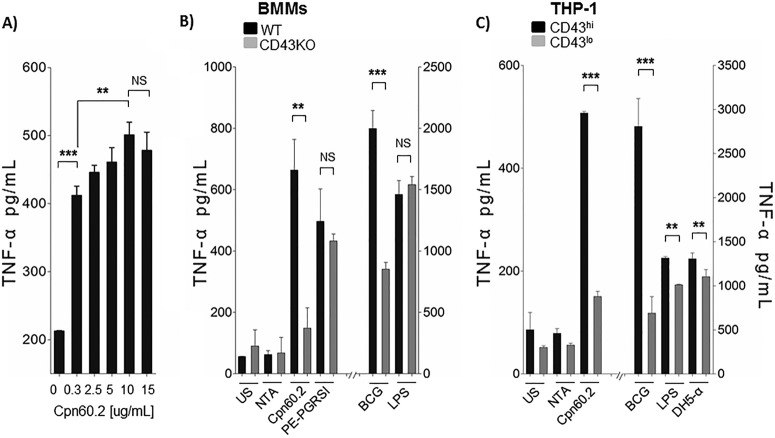
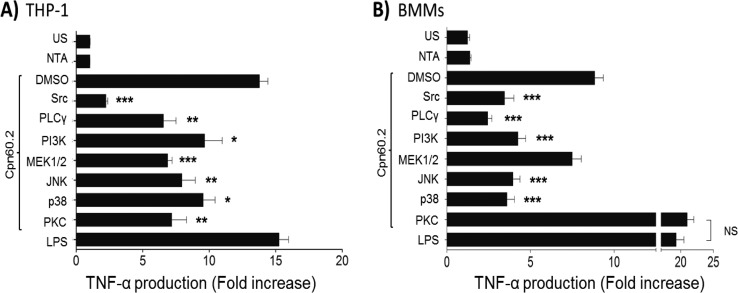
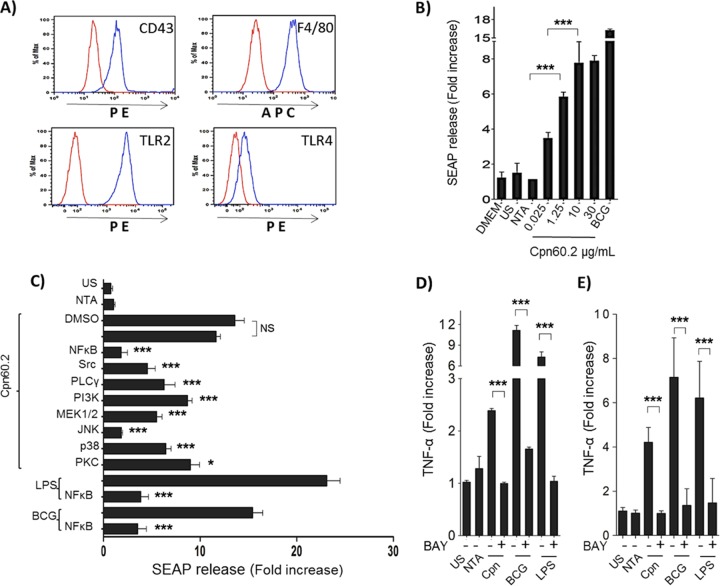
Mycobacterium tuberculosis is the causal agent of tuberculosis. Tumor necrosis factor alpha (TNF-α), transforming growth factor β (TGF-β), and gamma interferon (IFN-γ) secreted by activated macrophages and lymphocytes are considered essential to contain Mycobacterium tuberculosis infection. The CD43 sialomucin has been reported to act as a receptor for bacilli through its interaction with the chaperonin Cpn60.2, facilitating mycobacterium-macrophage contact. We report here that Cpn60.2 induces both human THP-1 cells and mouse-derived bone marrow-derived macrophages (BMMs) to produce TNF-α and that this production is CD43 dependent. In addition, we present evidence that the signaling pathway leading to TNF-α production upon interaction with Cpn60.2 requires active Src family kinases, phospholipase C-γ (PLC-γ), phosphatidylinositol 3-kinase (PI3K), p38, and Jun N-terminal protein kinase (JNK), both in BMMs and in THP-1 cells. Our data highlight the role of CD43 and Cpn60.2 in TNF-α production and underscore an important role for CD43 in the host-mycobacterium interaction.
Keywords: CD43; Cpn60.2; Mycobacterium tuberculosis; cell signaling; macrophages; tumor necrosis factor alpha.
Copyright © 2017 American Society for Microbiology.
Figures
Similar articles
-
Mycobacterium tuberculosis employs Cpn60.2 as an adhesin that binds CD43 on the macrophage surface.Cell Microbiol. 2010 Nov;12(11):1634-47. doi: 10.1111/j.1462-5822.2010.01496.x. Cell Microbiol. 2010. PMID: 20633027
-
CD43 controls the intracellular growth of Mycobacterium tuberculosis through the induction of TNF-alpha-mediated apoptosis.Cell Microbiol. 2008 Oct;10(10):2105-17. doi: 10.1111/j.1462-5822.2008.01194.x. Epub 2008 Jul 10. Cell Microbiol. 2008. PMID: 18637079
-
Mycobacterium tuberculosis Rv0652 stimulates production of tumour necrosis factor and monocytes chemoattractant protein-1 in macrophages through the Toll-like receptor 4 pathway.Immunology. 2012 Jun;136(2):231-40. doi: 10.1111/j.1365-2567.2012.03575.x. Immunology. 2012. PMID: 22385341 Free PMC article.
-
The rLrp of Mycobacterium tuberculosis inhibits proinflammatory cytokine production and downregulates APC function in mouse macrophages via a TLR2-mediated PI3K/Akt pathway activation-dependent mechanism.Cell Mol Immunol. 2016 Nov;13(6):729-746. doi: 10.1038/cmi.2015.58. Epub 2015 Jul 13. Cell Mol Immunol. 2016. PMID: 26166760 Free PMC article.
-
Regulation of the MIR155 host gene in physiological and pathological processes.Gene. 2013 Dec 10;532(1):1-12. doi: 10.1016/j.gene.2012.12.009. Epub 2012 Dec 14. Gene. 2013. PMID: 23246696 Review.
Cited by
-
Rv2346c enhances mycobacterial survival within macrophages by inhibiting TNF-α and IL-6 production via the p38/miRNA/NF-κB pathway.Emerg Microbes Infect. 2018 Sep 19;7(1):158. doi: 10.1038/s41426-018-0162-6. Emerg Microbes Infect. 2018. PMID: 30232332 Free PMC article.
-
PPE38-Secretion-Dependent Proteins of M. tuberculosis Alter NF-kB Signalling and Inflammatory Responses in Macrophages.Front Immunol. 2021 Jul 2;12:702359. doi: 10.3389/fimmu.2021.702359. eCollection 2021. Front Immunol. 2021. PMID: 34276695 Free PMC article.
-
Negative Regulation of Autophagy during Macrophage Infection by Mycobacterium bovis BCG via Protein Kinase C Activation.Int J Mol Sci. 2024 Mar 9;25(6):3145. doi: 10.3390/ijms25063145. Int J Mol Sci. 2024. PMID: 38542119 Free PMC article.
References
-
- Gaynor CD, McCormack FX, Voelker DR, McGowan SE, Schlesinger LS. 1995. Pulmonary surfactant protein A mediates enhanced phagocytosis of Mycobacterium tuberculosis by a direct interaction with human macrophages. J Immunol 155:5343–5351. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
