

Dual role of ALCAM in neuroinflammation and blood-brain barrier homeostasis
- PMID: 28069965
- PMCID: PMC5278491
- DOI: 10.1073/pnas.1614336114
Dual role of ALCAM in neuroinflammation and blood-brain barrier homeostasis
Abstract
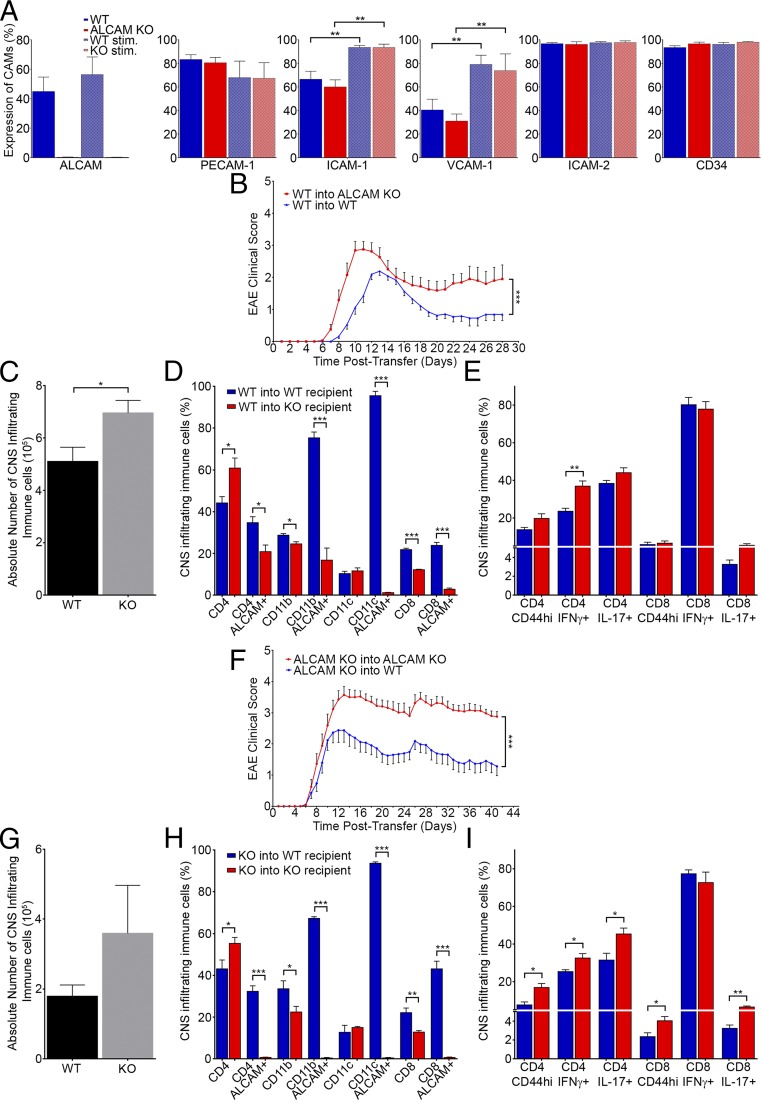
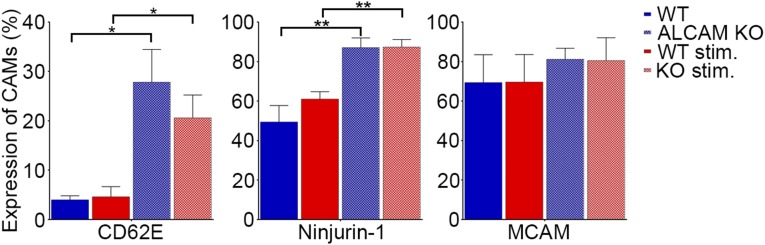
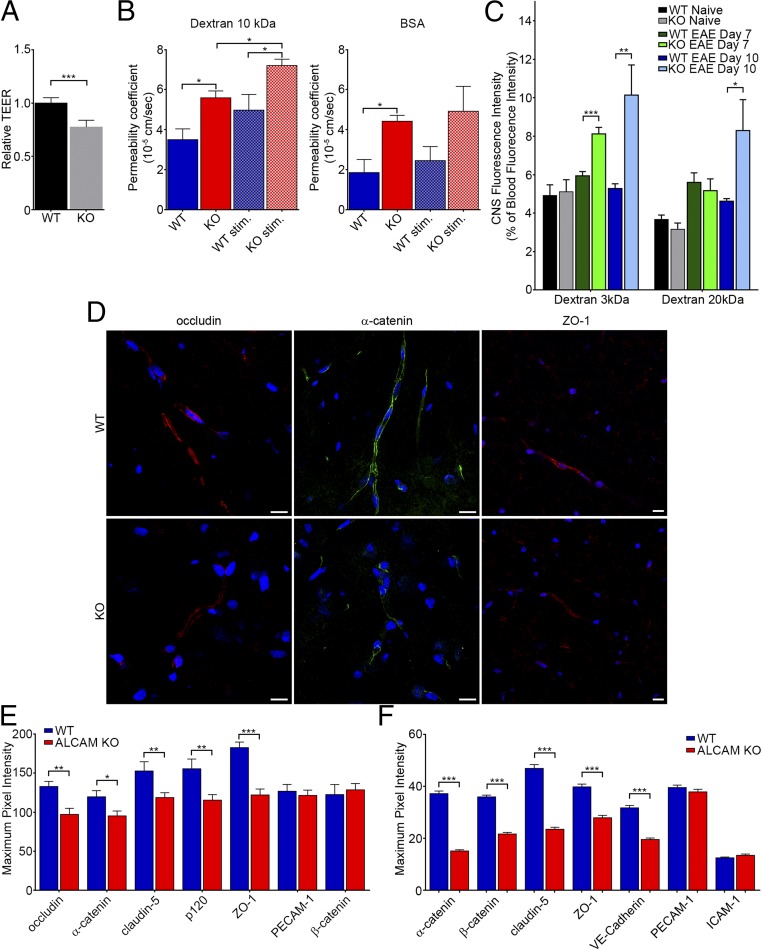
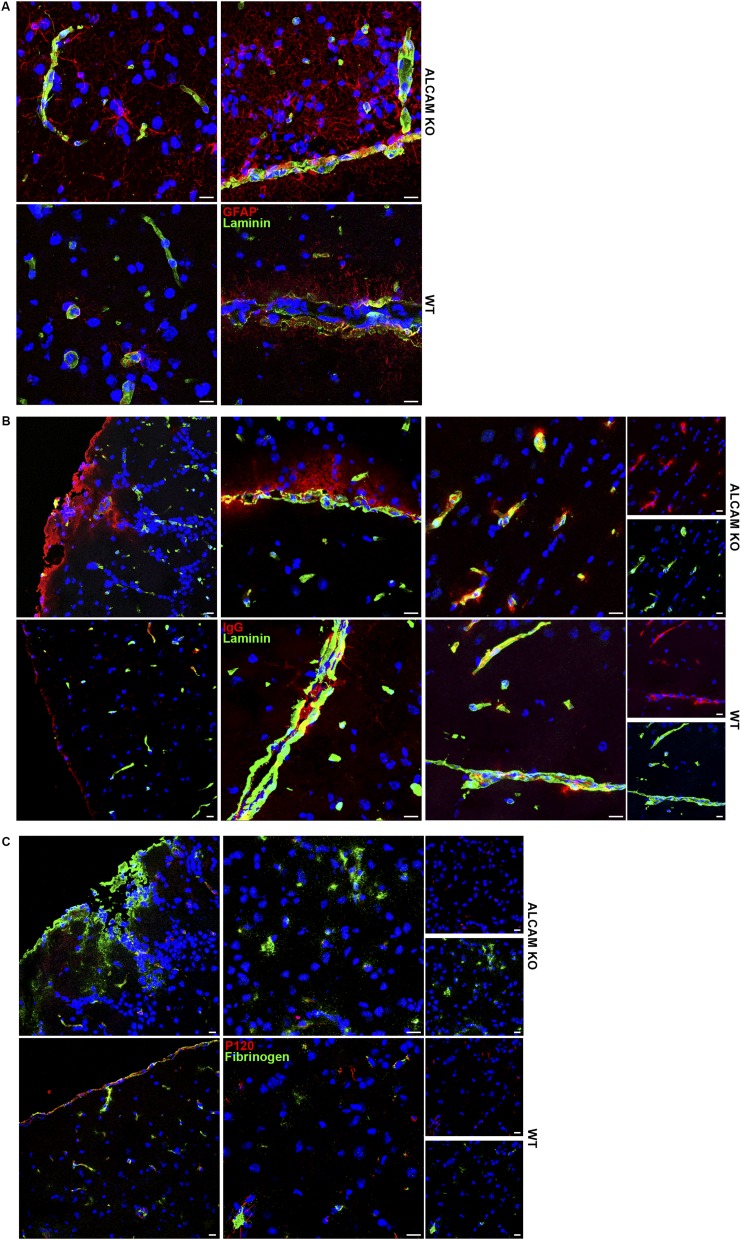
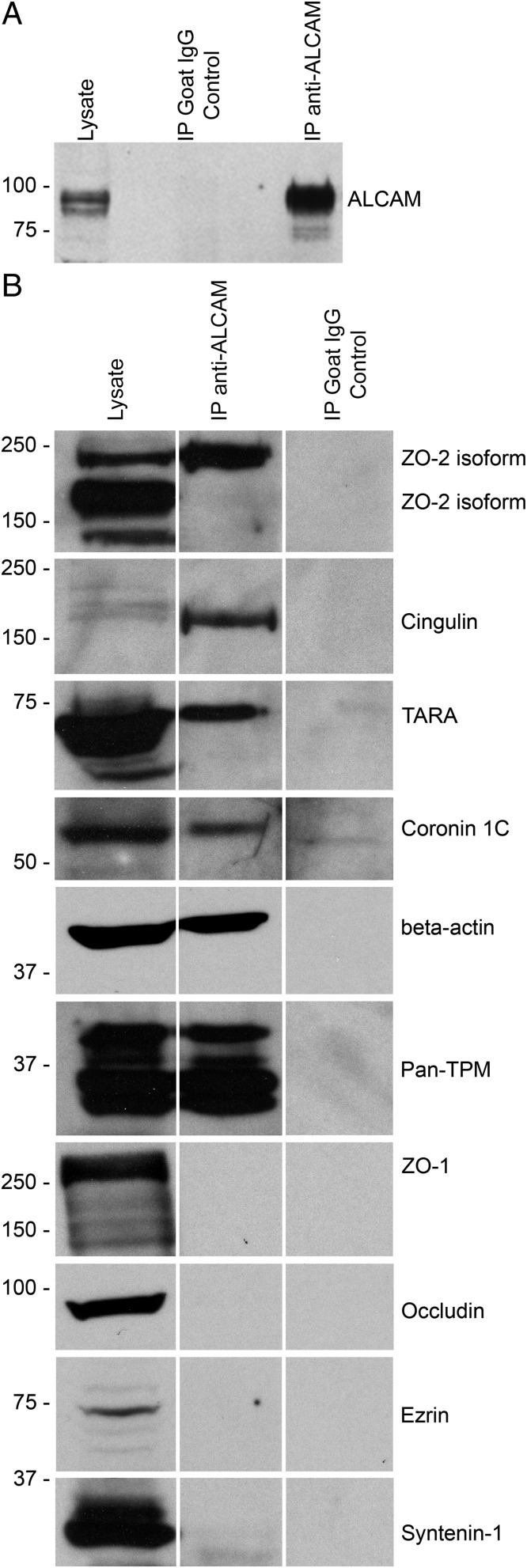
Activated leukocyte cell adhesion molecule (ALCAM) is a cell adhesion molecule found on blood-brain barrier endothelial cells (BBB-ECs) that was previously shown to be involved in leukocyte transmigration across the endothelium. In the present study, we found that ALCAM knockout (KO) mice developed a more severe myelin oligodendrocyte glycoprotein (MOG)35-55-induced experimental autoimmune encephalomyelitis (EAE). The exacerbated disease was associated with a significant increase in the number of CNS-infiltrating proinflammatory leukocytes compared with WT controls. Passive EAE transfer experiments suggested that the pathophysiology observed in active EAE was linked to the absence of ALCAM on BBB-ECs. In addition, phenotypic characterization of unimmunized ALCAM KO mice revealed a reduced expression of BBB junctional proteins. Further in vivo, in vitro, and molecular analysis confirmed that ALCAM is associated with tight junction molecule assembly at the BBB, explaining the increased permeability of CNS blood vessels in ALCAM KO animals. Collectively, our data point to a biologically important function of ALCAM in maintaining BBB integrity.
Keywords: ALCAM; EAE; blood–brain barrier; multiple sclerosis; tight junctions.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
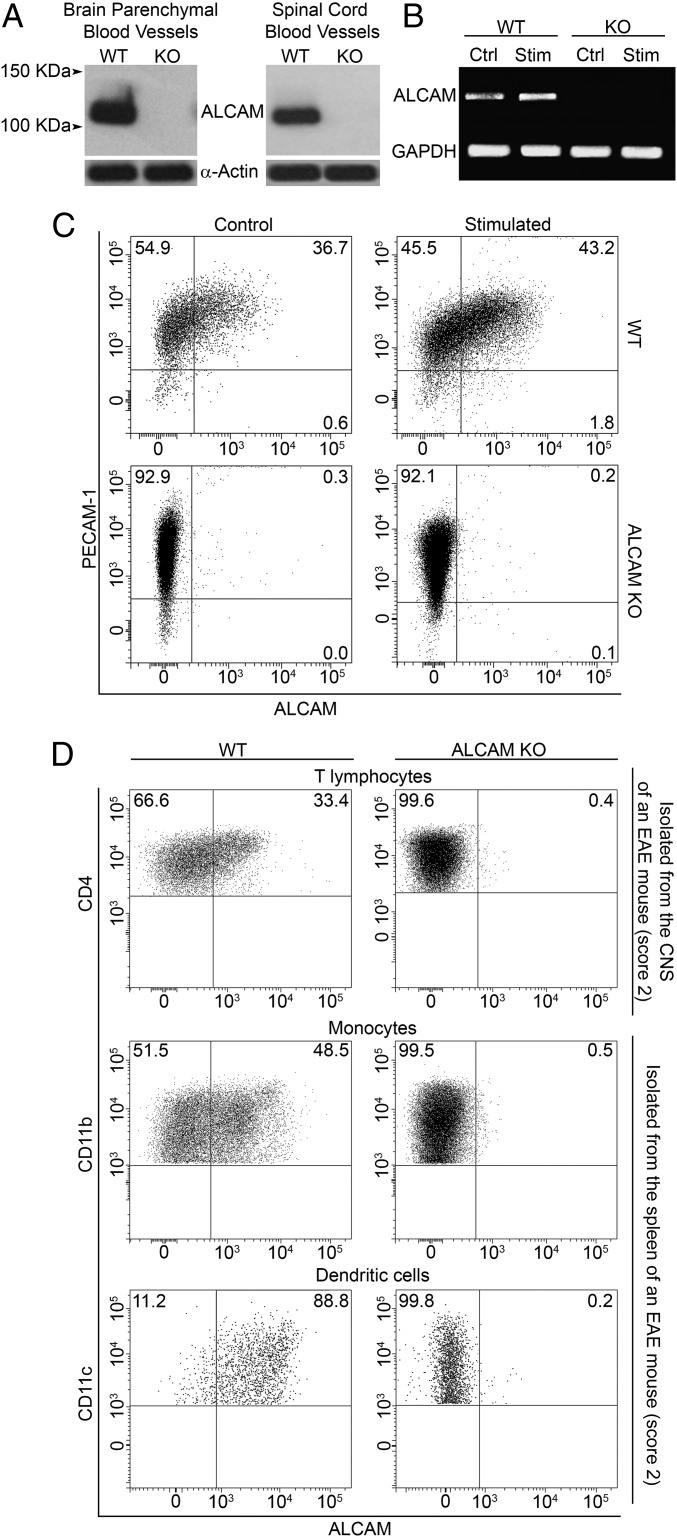
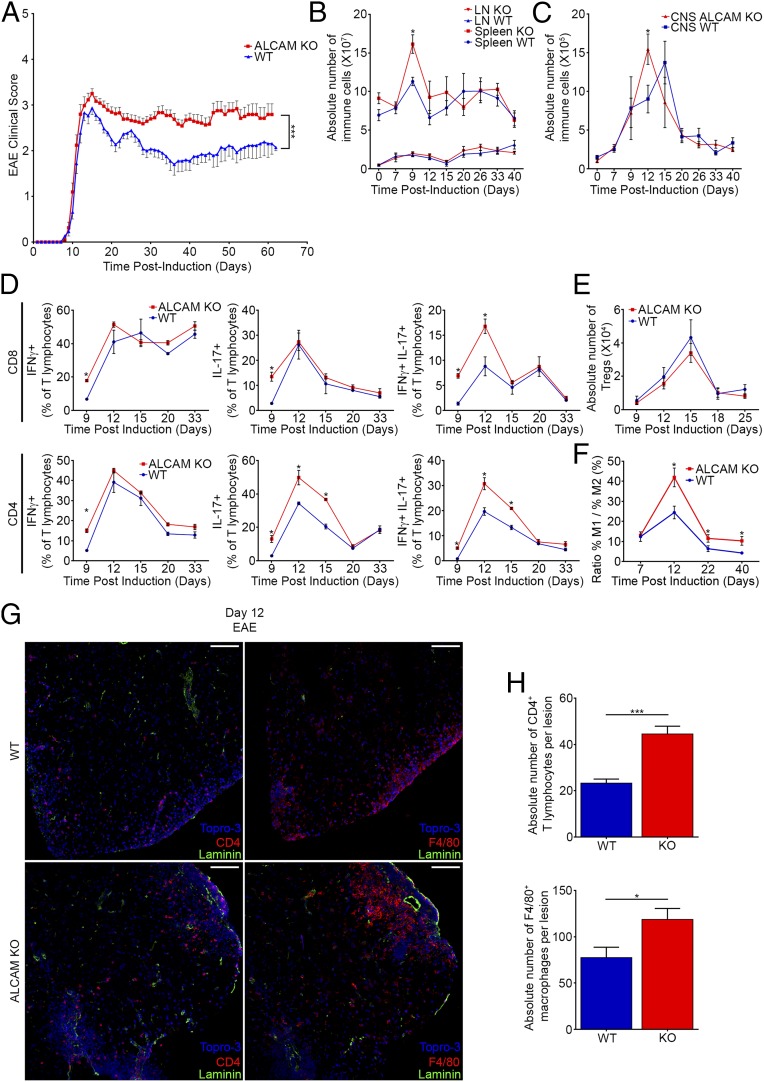
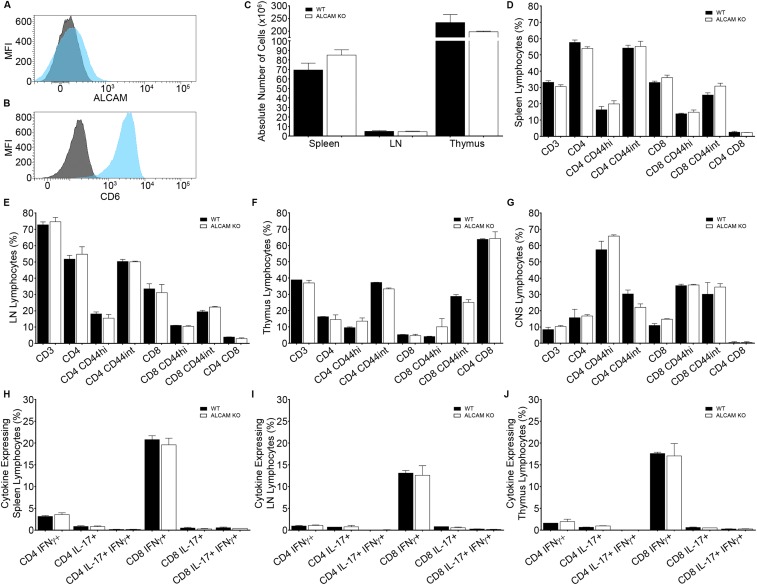
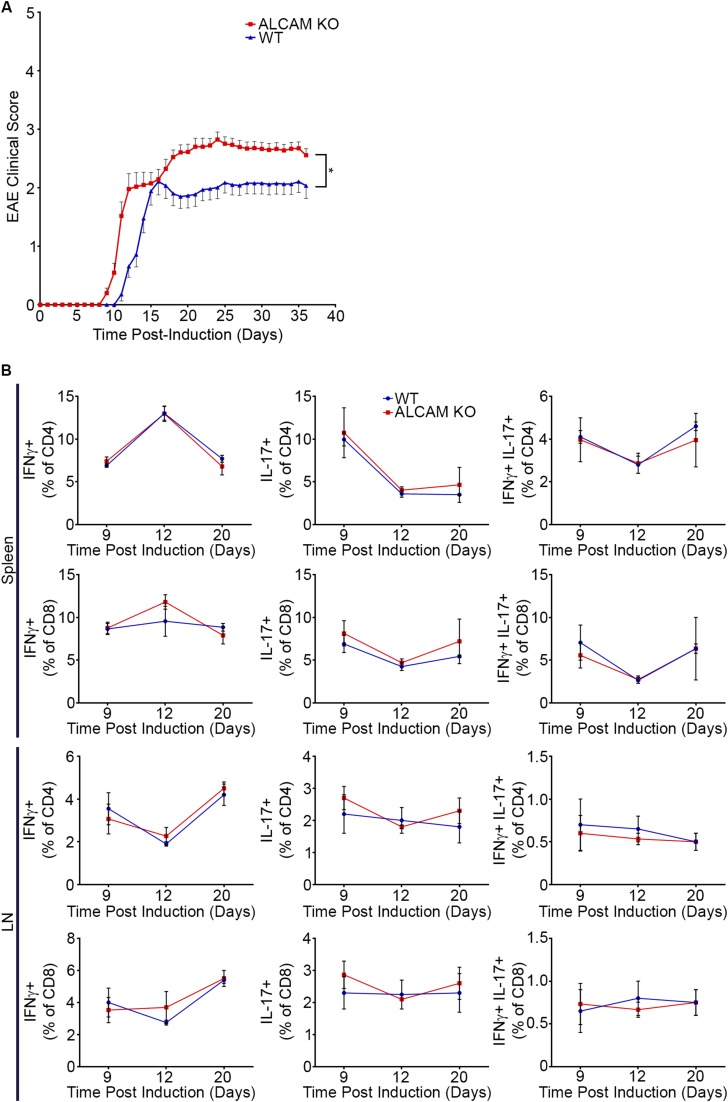
References
-
- Cayrol R, Haqqani AS, Ifergan I, Dodelet-Devillers A, Prat A. Isolation of human brain endothelial cells and characterization of lipid raft-associated proteins by mass spectroscopy. Methods Mol Biol. 2011;686:275–295. - PubMed
-
- Dodelet-Devillers A, et al. Functions of lipid raft membrane microdomains at the blood-brain barrier. J Mol Med (Berl) 2009;87(8):765–774. - PubMed
-
- Larochelle C, Alvarez JI, Prat A. How do immune cells overcome the blood-brain barrier in multiple sclerosis? FEBS Lett. 2011;585(23):3770–3780. - PubMed
-
- Alvarez JI, Cayrol R, Prat A. Disruption of central nervous system barriers in multiple sclerosis. Biochim Biophys Acta. 2011;1812(2):252–264. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
