

spict, a cyst cell-specific gene, regulates starvation-induced spermatogonial cell death in the Drosophila testis
- PMID: 28071722
- PMCID: PMC5223112
- DOI: 10.1038/srep40245
spict, a cyst cell-specific gene, regulates starvation-induced spermatogonial cell death in the Drosophila testis
Abstract
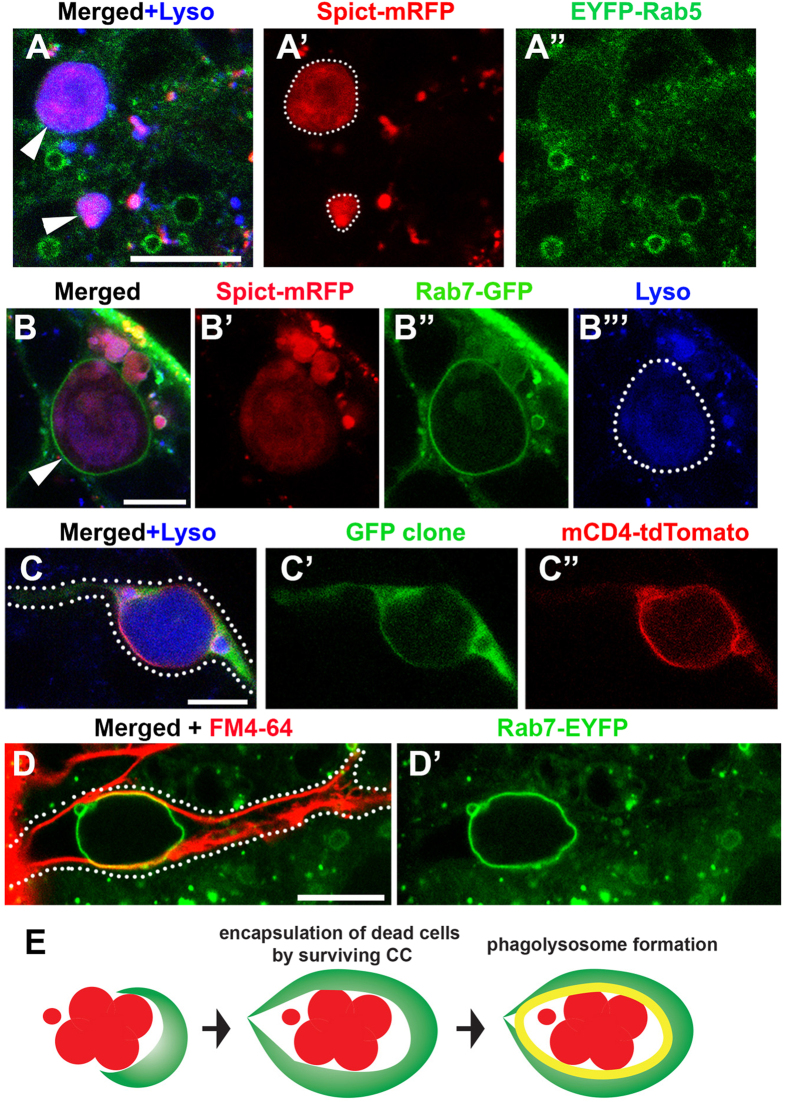
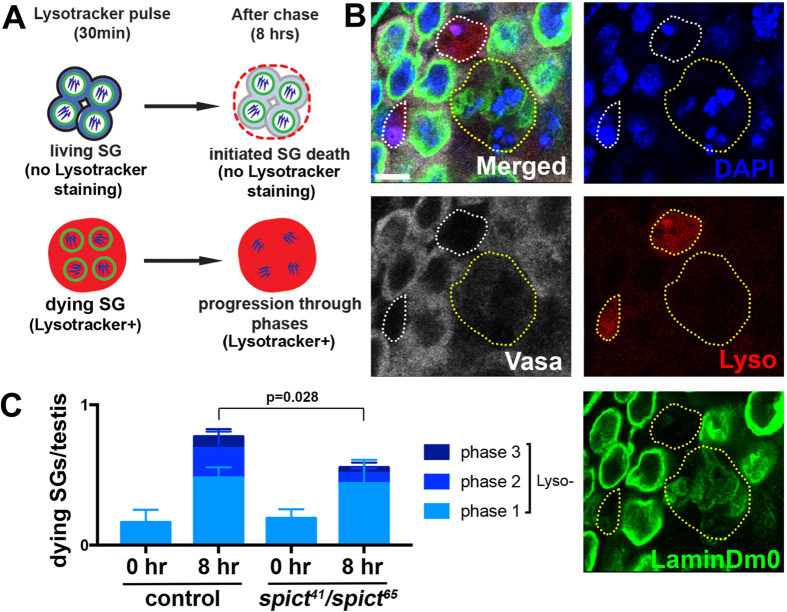
Tissues are maintained in a homeostatic state by balancing the constant loss of old cells with the continued production of new cells. Tissue homeostasis can shift between high and low turnover states to cope with environmental changes such as nutrient availability. Recently, we discovered that the elimination of transit-amplifying cells plays a critical role in maintaining the stem cell population during protein starvation in the Drosophila testis. Here, we identify spict, a gene expressed specifically in differentiating cyst cells, as a regulator of spermatogonial death. Spict is upregulated in cyst cells that phagocytose dying spermatogonia. We propose that phagocytosis and subsequent clearance of dead spermatogonia, which is partly promoted by Spict, contribute to stem cell maintenance during prolonged protein starvation.
Figures
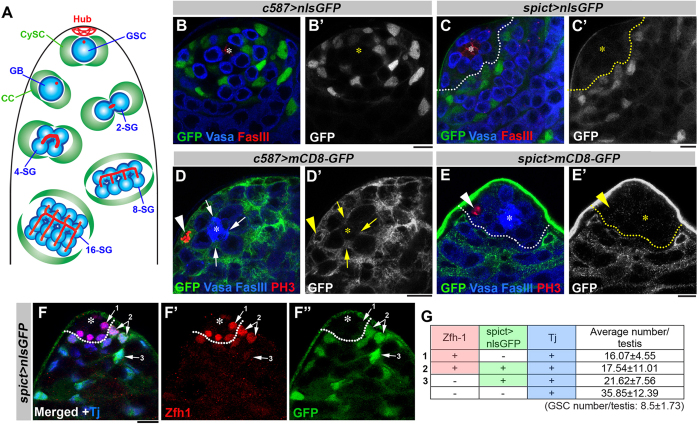
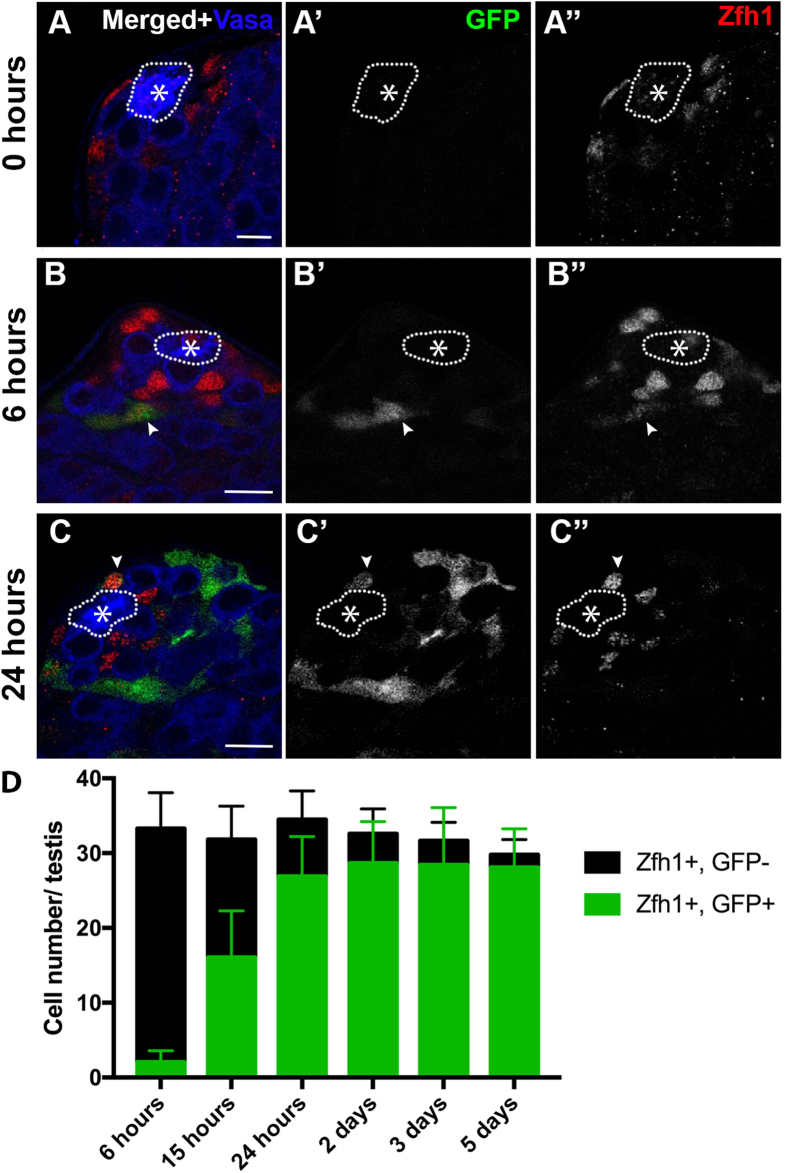
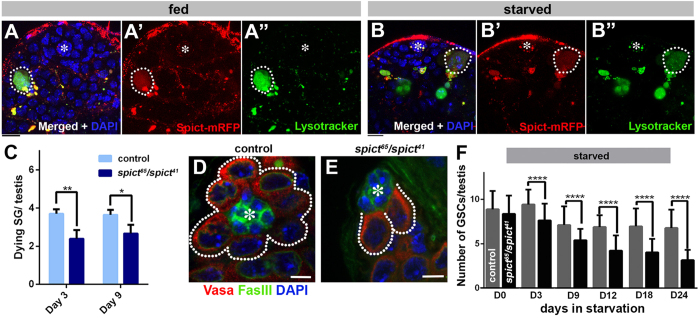
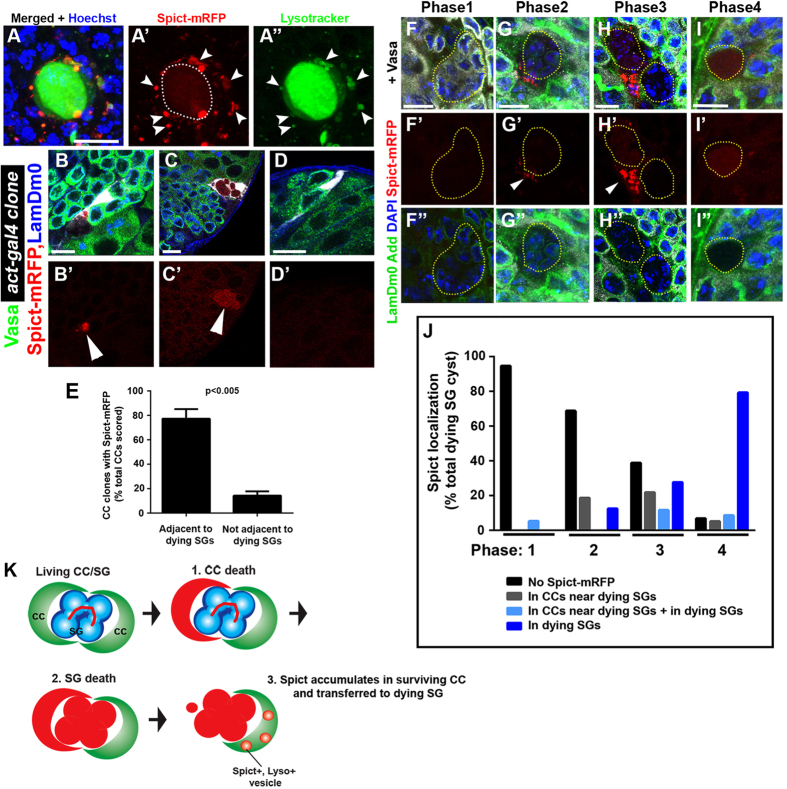
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
