

Nup82 functions redundantly with Nup136 in a salicylic acid-dependent defense response of Arabidopsis thaliana
- PMID: 28071978
- PMCID: PMC5499915
- DOI: 10.1080/19491034.2017.1279774
Nup82 functions redundantly with Nup136 in a salicylic acid-dependent defense response of Arabidopsis thaliana
Abstract
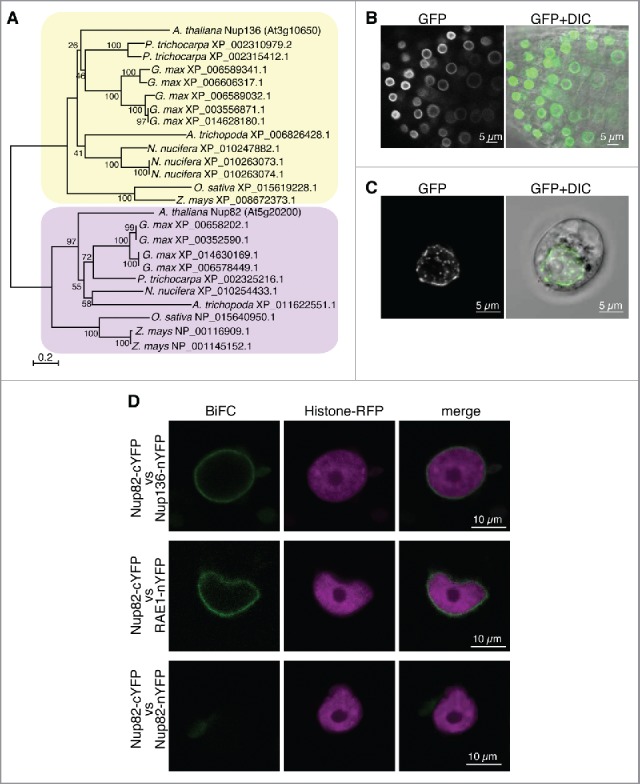
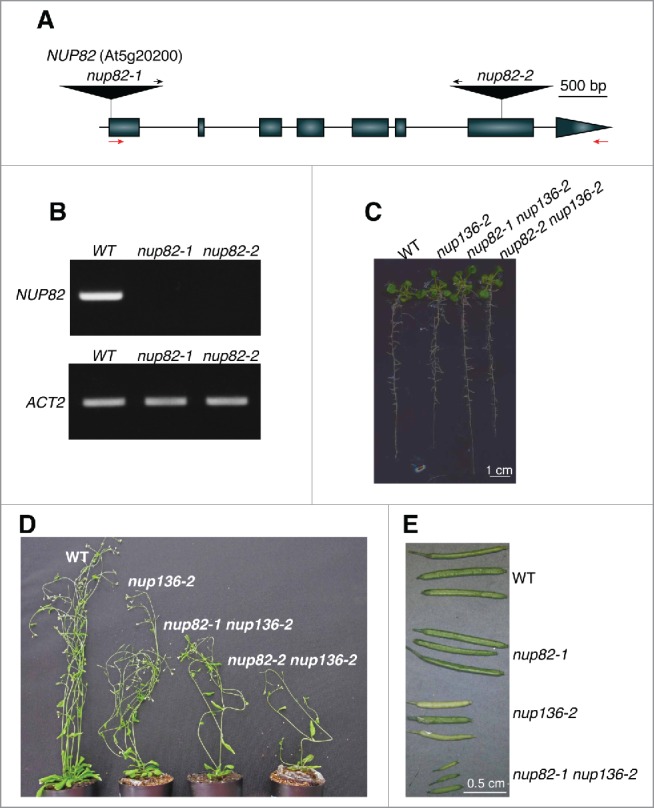
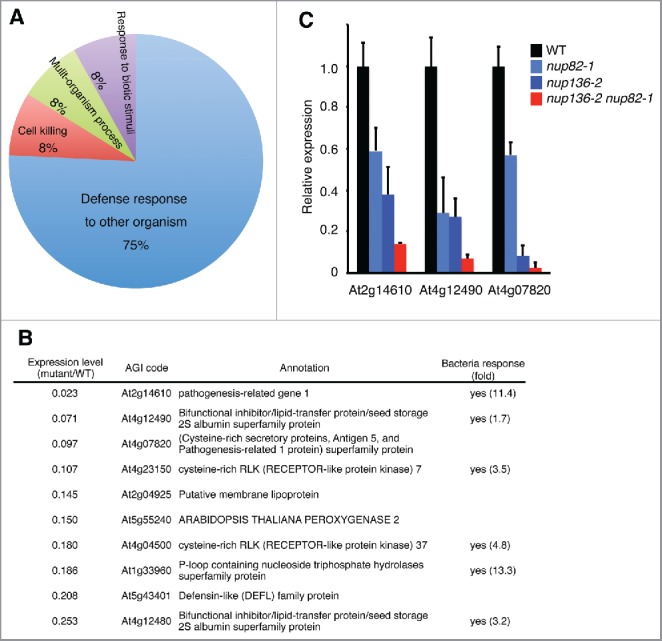
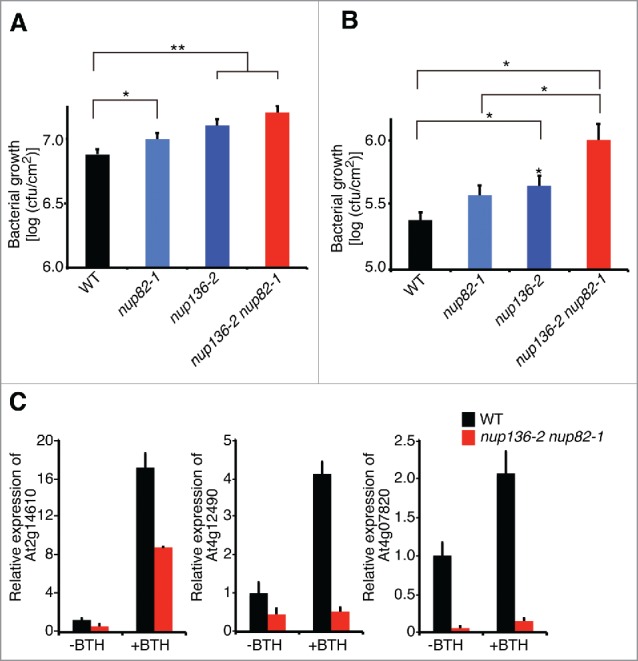
The nuclear pore complex (NPC) comprises more than 30 nucleoporins (Nups). NPC mediates macromolecular trafficking between the nucleoplasm and the cytoplasm, but specific roles of individual Nups are poorly understood in higher plants. Here, we show that the novel nucleoporin unique to angiosperm plants (designated as Nup82) functions in a salicylic acid-dependent defense in a redundant manner with Nup136, which is a component of the nuclear basket in the NPC. Arabidopsis thaliana Nup82 had a similar amino acid sequence to the N-terminal half of Nup136 and a Nup82-GFP fusion was localized on the nuclear envelope. Immunoprecipitation and bimolecular fluorescence complementation analyses revealed that Nup82 interacts with the NPC components Nup136 and RAE1. The double knockout mutant nup82 nup136 showed severe growth defects, while the single knockout mutant nup82 did not, suggesting that Nup82 functions redundantly with Nup136. nup82 nup136 impaired benzothiadiazole (an analog of salicylic acid)-induced resistance to the virulent bacteria Pseudomonas syringae pv. tomato DC3000. Furthermore, transcriptome analysis of nup82 nup136 indicates that deficiency of Nup82 and Nup136 causes noticeable downregulation of immune-related genes. These results suggest that Nup82 and Nup136 are redundantly involved in transcriptional regulation of salicylic acid-responsive genes through nuclear transport of signaling molecules.
Keywords: Arabidopsis thaliana; Nup136/Nup1; Nup82; nuclear envelope; nucleoporin; plant immunity; proteome; transcriptome.
Figures
References
-
- Speese SD, Ashley J, Jokhi V, Nunnari J, Barria R, Li Y, Ataman B, Koon A, Chang YT, Li Q, et al.. Nuclear envelope budding enables large ribonucleoprotein particle export during synaptic Wnt signaling. Cell 2012; 149:832-46; PMID:22579286; https://doi.org/ 10.1016/j.cell.2012.03.032 - DOI - PMC - PubMed
-
- Roberts K, Northcote DH. Structure of the nuclear pore in higher plants. Nature 1970; 228:385-6; PMID:5473989; https://doi.org/ 10.1038/228385a0 - DOI - PubMed
-
- Fiserova J, Goldberg MW. Relationships at the nuclear envelope: lamins and nuclear pore complexes in animals and plants. Biochem Soc Trans 2010; 38:829-31; PMID:20491671; https://doi.org/ 10.1042/BST0380829 - DOI - PubMed
-
- Fiserova J, Kiseleva E, Goldberg MW. Nuclear envelope and nuclear pore complex structure and organization in tobacco BY-2 cells. Plant J 2009; 59:243-55; PMID:19392704; https://doi.org/ 10.1111/j.1365-313X.2009.03865.x - DOI - PubMed
-
- Rout MP, Aitchison JD, Suprapto A, Hjertaas K, Zhao Y, Chait BT. The yeast nuclear pore complex: composition, architecture, and transport mechanism. J Cell Biol 2000; 148:635-51; PMID:10684247; https://doi.org/ 10.1083/jcb.148.4.635 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases