

Dopaminergic dynamics underlying sex-specific cocaine reward
- PMID: 28072417
- PMCID: PMC5234081
- DOI: 10.1038/ncomms13877
Dopaminergic dynamics underlying sex-specific cocaine reward
Abstract
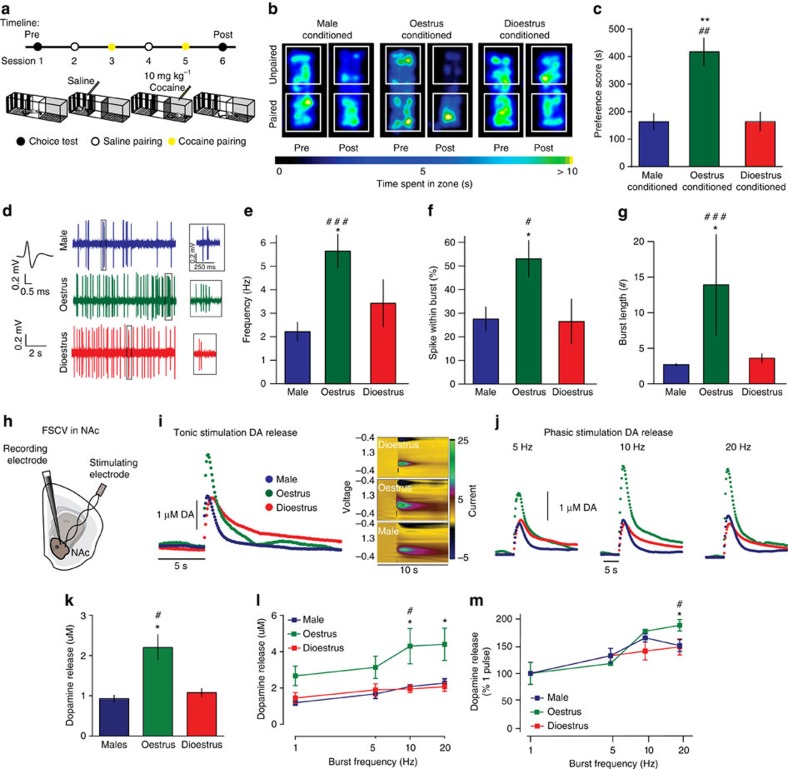
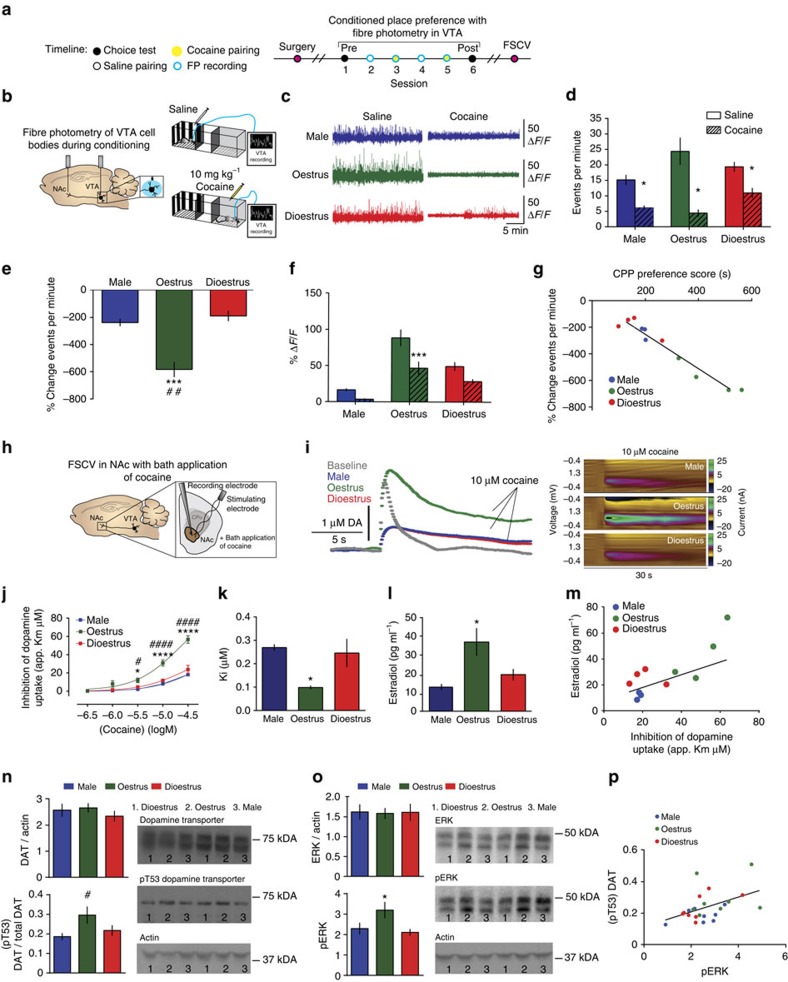
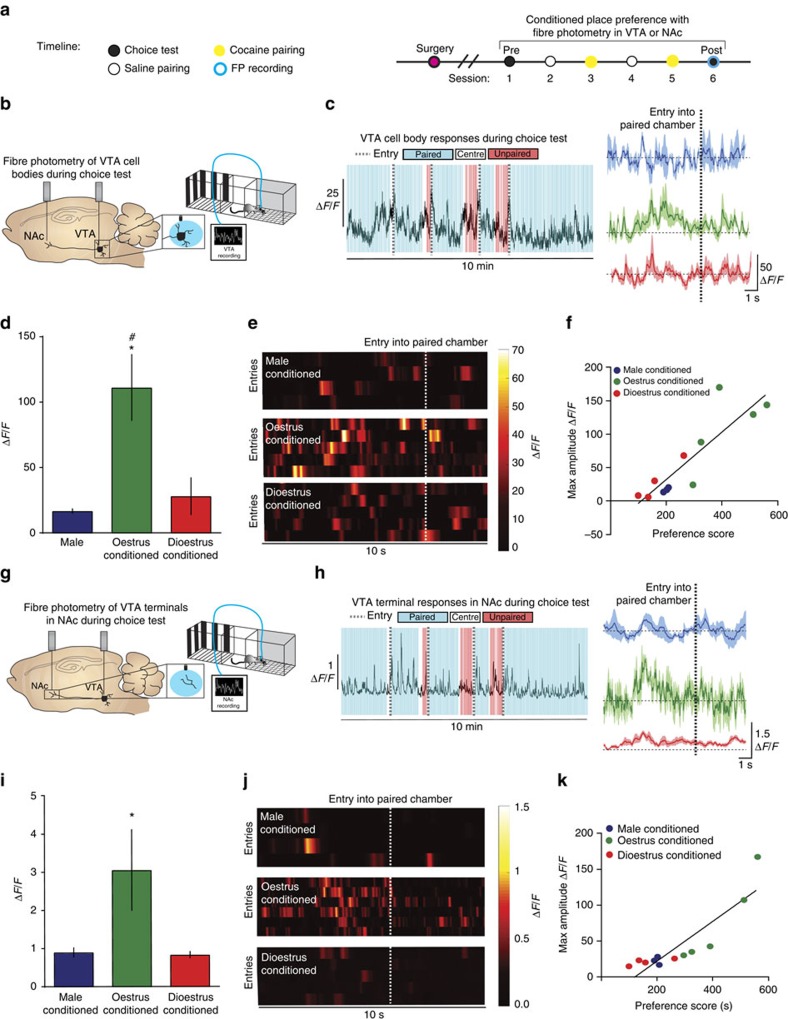
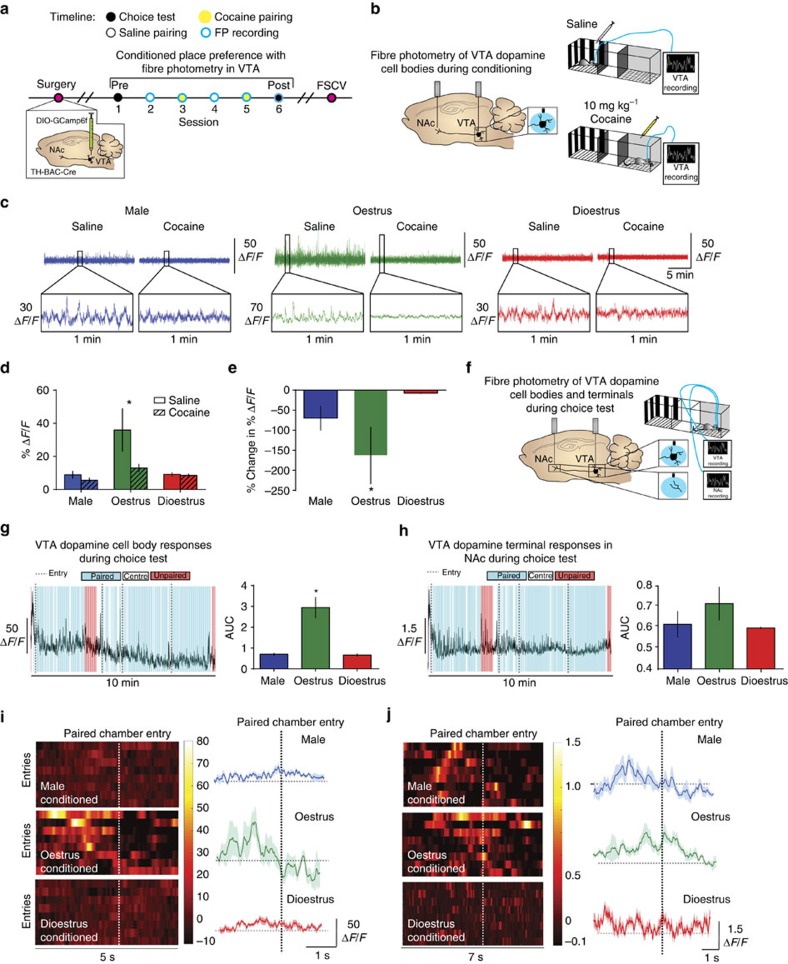
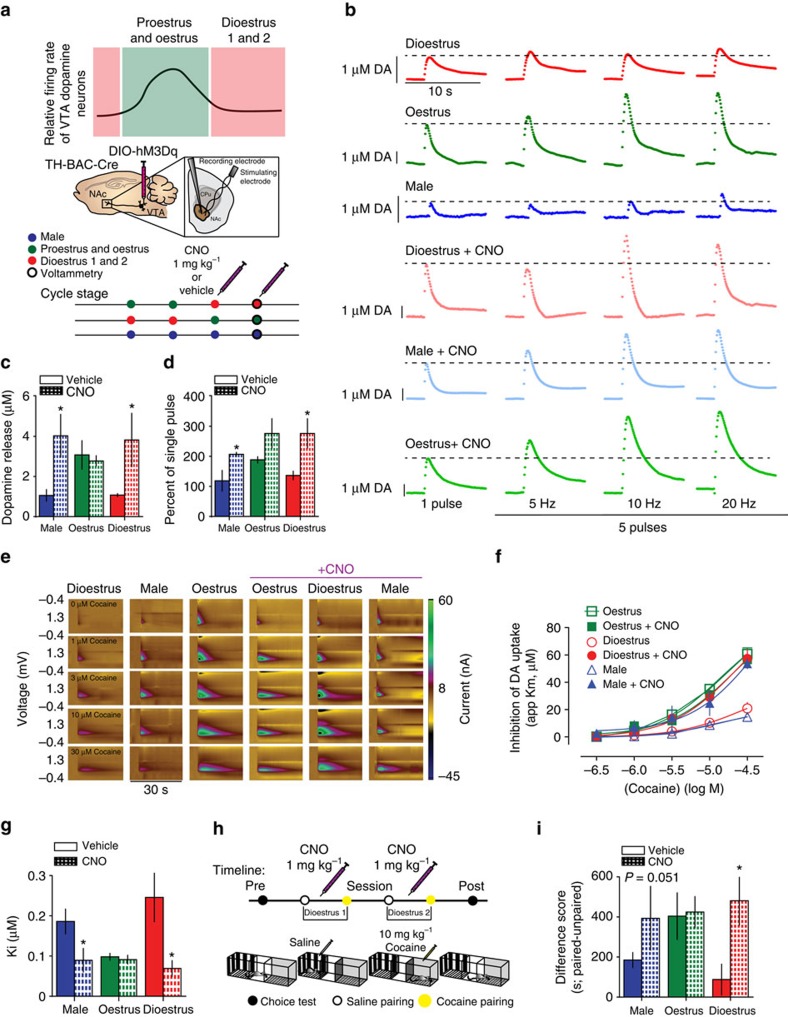
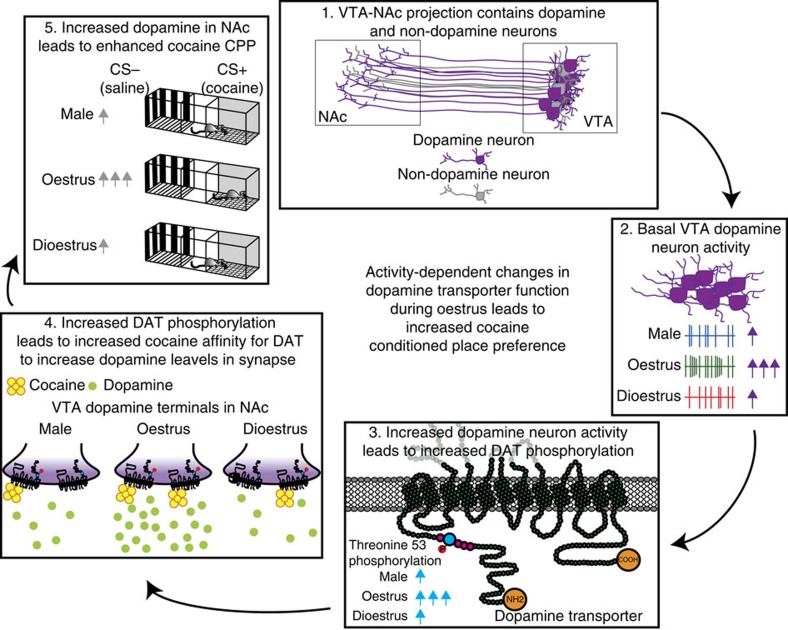
Although both males and females become addicted to cocaine, females transition to addiction faster and experience greater difficulties remaining abstinent. We demonstrate an oestrous cycle-dependent mechanism controlling increased cocaine reward in females. During oestrus, ventral tegmental area (VTA) dopamine neuron activity is enhanced and drives post translational modifications at the dopamine transporter (DAT) to increase the ability of cocaine to inhibit its function, an effect mediated by estradiol. Female mice conditioned to associate cocaine with contextual cues during oestrus have enhanced mesolimbic responses to these cues in the absence of drug. Using chemogenetic approaches, we increase VTA activity to mechanistically link oestrous cycle-dependent enhancement of VTA firing to enhanced cocaine affinity at DAT and subsequent reward processing. These data have implications for sexual dimorphism in addiction vulnerability and define a mechanism by which cellular activity results in protein alterations that contribute to dysfunctional learning and reward processing.
Figures
References
-
- Reid A. G., Lingford-Hughes A. R., Cancela L. M. & Kalivas P. W. Substance abuse disorders. Handb. Clin. Neurol. 106, 419–431 (2012). - PubMed
-
- Van Etten M. L., Neumark Y. D. & Anthony J. C. Male-female differences in the earliest stages of drug involvement. Addiction 94, 1413–1419 (1999). - PubMed
-
- Randall C. L. et al. Telescoping of landmark events associated with drinking: a gender comparison. J. Stud. Alcohol 60, 252–260 (1999). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
