

Overexpression of PGC-1α increases peroxisomal activity and mitochondrial fatty acid oxidation in human primary myotubes
- PMID: 28073778
- PMCID: PMC5406987
- DOI: 10.1152/ajpendo.00331.2016
Overexpression of PGC-1α increases peroxisomal activity and mitochondrial fatty acid oxidation in human primary myotubes
Abstract
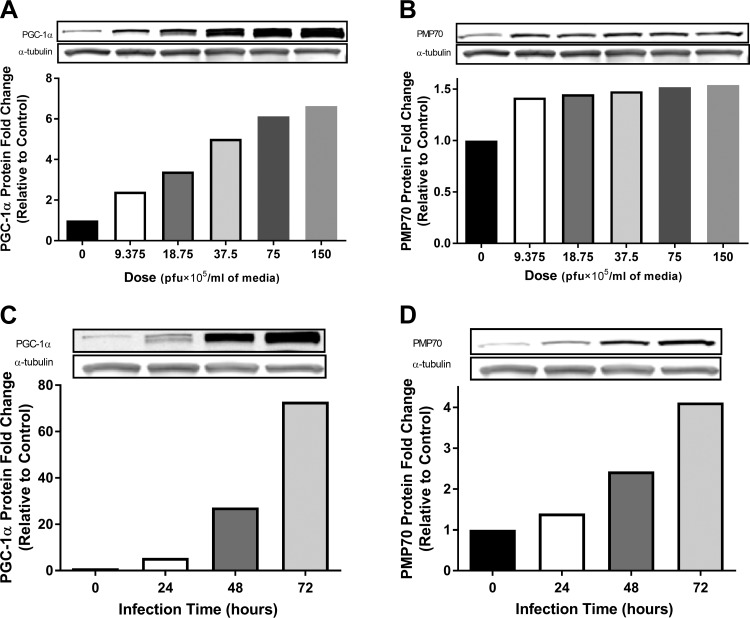
Peroxisomes are indispensable organelles for lipid metabolism in humans, and their biogenesis has been assumed to be under regulation by peroxisome proliferator-activated receptors (PPARs). However, recent studies in hepatocytes suggest that the mitochondrial proliferator PGC-1α (peroxisome proliferator-activated receptor gamma coactivator-1α) also acts as an upstream transcriptional regulator for enhancing peroxisomal abundance and associated activity. It is unknown whether the regulatory mechanism(s) for enhancing peroxisomal function is through the same node as mitochondrial biogenesis in human skeletal muscle (HSkM) and whether fatty acid oxidation (FAO) is affected. Primary myotubes from vastus lateralis biopsies from lean donors (BMI = 24.0 ± 0.6 kg/m2; n = 6) were exposed to adenovirus encoding human PGC-1α or GFP control. Peroxisomal biogenesis proteins (peroxins) and genes (PEXs) responsible for proliferation and functions were assessed by Western blotting and real-time qRT-PCR, respectively. [1-14C]palmitic acid and [1-14C]lignoceric acid (exclusive peroxisomal-specific substrate) were used to assess mitochondrial oxidation of peroxisomal-derived metabolites. After overexpression of PGC-1α, 1) peroxisomal membrane protein 70 kDa (PMP70), PEX19, and mitochondrial citrate synthetase protein content were significantly elevated (P < 0.05), 2) PGC-1α, PMP70, key PEXs, and peroxisomal β-oxidation mRNA expression levels were significantly upregulated (P < 0.05), and 3) a concomitant increase in lignoceric acid oxidation by both peroxisomal and mitochondrial activity was observed (P < 0.05). These novel findings demonstrate that, in addition to the proliferative effect on mitochondria, PGC-1α can induce peroxisomal activity and accompanying elevations in long-chain and very-long-chain fatty acid oxidation by a peroxisomal-mitochondrial functional cooperation, as observed in HSkM cells.
Keywords: human skeletal muscle cells; lignoceric acid oxidation; obesity; peroxisome proliferator-activated receptors; β-oxidation.
Copyright © 2017 the American Physiological Society.
Figures




References
-
- Amati F, Dubé JJ, Alvarez-Carnero E, Edreira MM, Chomentowski P, Coen PM, Switzer GE, Bickel PE, Stefanovic-Racic M, Toledo FGS, Goodpaster BH. Skeletal muscle triglycerides, diacylglycerols, and ceramides in insulin resistance: another paradox in endurance-trained athletes? Diabetes 60: 2588–2597, 2011. doi: 10.2337/db10-1221. - DOI - PMC - PubMed
-
- Anderson EJ, Lustig ME, Boyle KE, Woodlief TL, Kane DA, Lin CT, Price JW III, Kang L, Rabinovitch PS, Szeto HH, Houmard JA, Cortright RN, Wasserman DH, Neufer PD. Mitochondrial H2O2 emission and cellular redox state link excess fat intake to insulin resistance in both rodents and humans. J Clin Invest 119: 573–581, 2009. doi: 10.1172/JCI37048. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous