

Evidence of a Vocalic Proto-System in the Baboon (Papio papio) Suggests Pre-Hominin Speech Precursors
- PMID: 28076426
- PMCID: PMC5226677
- DOI: 10.1371/journal.pone.0169321
Evidence of a Vocalic Proto-System in the Baboon (Papio papio) Suggests Pre-Hominin Speech Precursors
Abstract
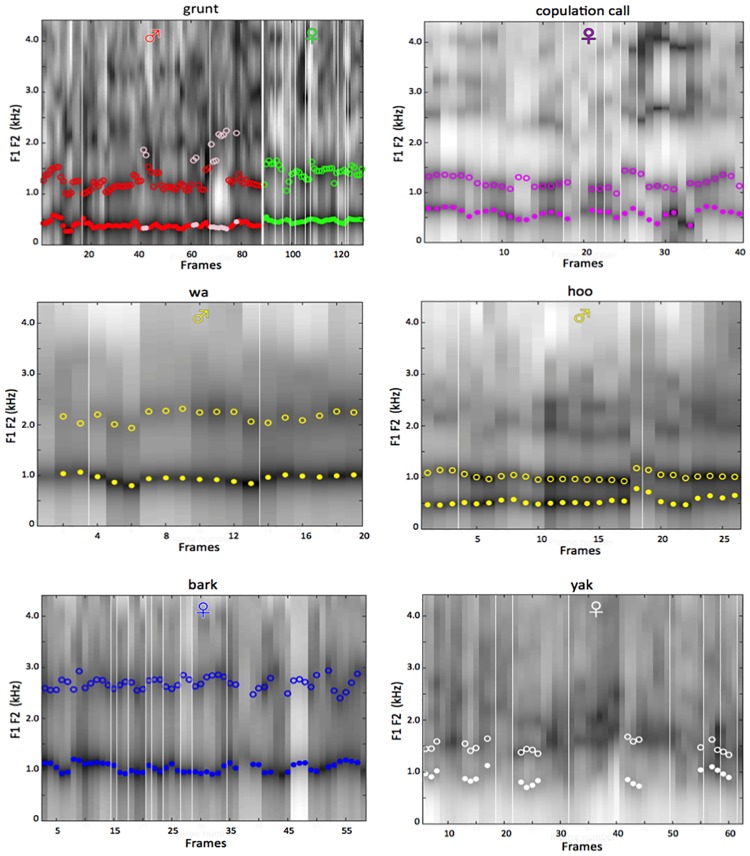
Language is a distinguishing characteristic of our species, and the course of its evolution is one of the hardest problems in science. It has long been generally considered that human speech requires a low larynx, and that the high larynx of nonhuman primates should preclude their producing the vowel systems universally found in human language. Examining the vocalizations through acoustic analyses, tongue anatomy, and modeling of acoustic potential, we found that baboons (Papio papio) produce sounds sharing the F1/F2 formant structure of the human [ɨ æ ɑ ɔ u] vowels, and that similarly with humans those vocalic qualities are organized as a system on two acoustic-anatomic axes. This confirms that hominoids can produce contrasting vowel qualities despite a high larynx. It suggests that spoken languages evolved from ancient articulatory skills already present in our last common ancestor with Cercopithecoidea, about 25 MYA.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




References
-
- International Phonetic Association. Handbook of the International Phonetic Association: a guide to the use of the International Phonetic Alphabet. Cambridge, U.K.; New York, NY: Cambridge University Press; 1999.
-
- Maddieson I. Patterns of sounds. Cambridge, UK; New York: Cambridge University Press; 2009.
-
- Lieberman PH. The evolution of human speech: its anatomical and neural bases. Curr Anthropol. 2007;48: 39–66.
-
- Lieberman PH, Klatt DH, Wilson WH. Vocal tract limitations on the vowel repertoires of rhesus monkey and other nonhuman primates. Science. 1969;164: 1185–1187. - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
