

Coupled Proliferation and Apoptosis Maintain the Rapid Turnover of Microglia in the Adult Brain
- PMID: 28076784
- PMCID: PMC5263237
- DOI: 10.1016/j.celrep.2016.12.041
Coupled Proliferation and Apoptosis Maintain the Rapid Turnover of Microglia in the Adult Brain
Abstract
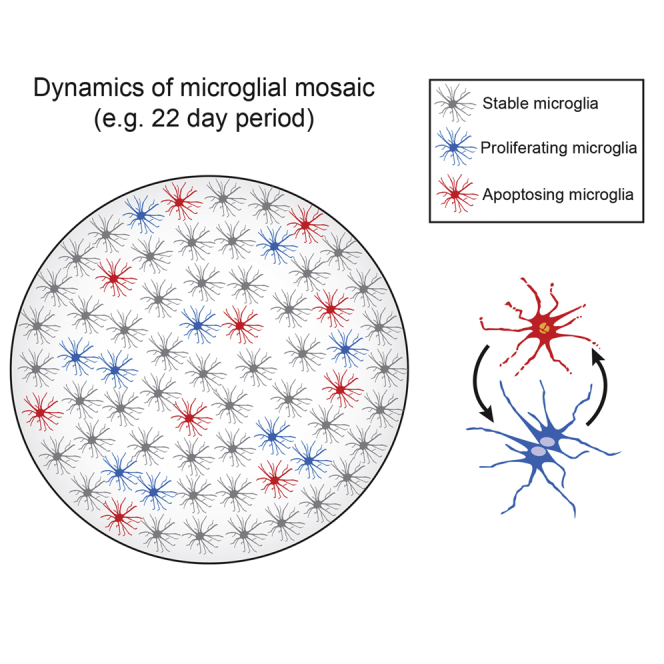
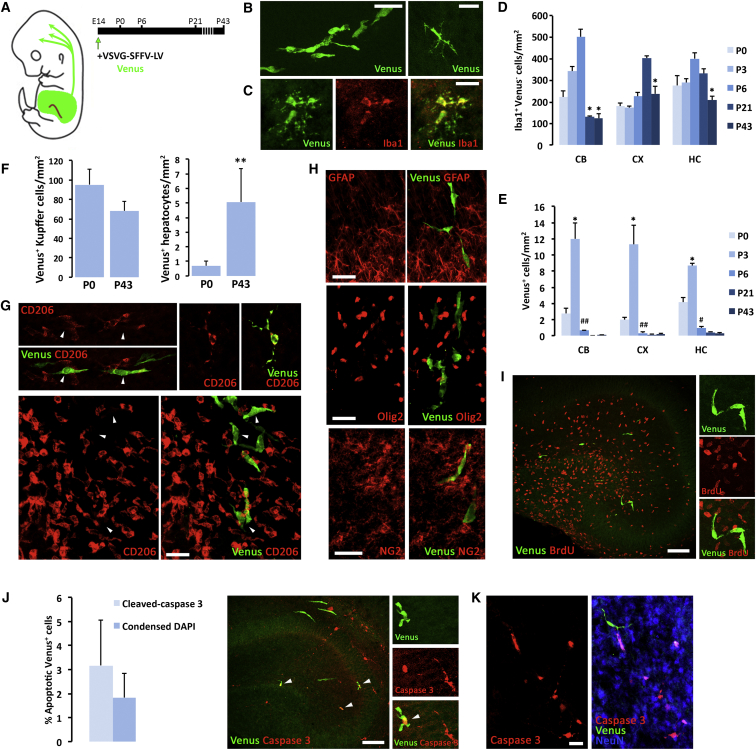
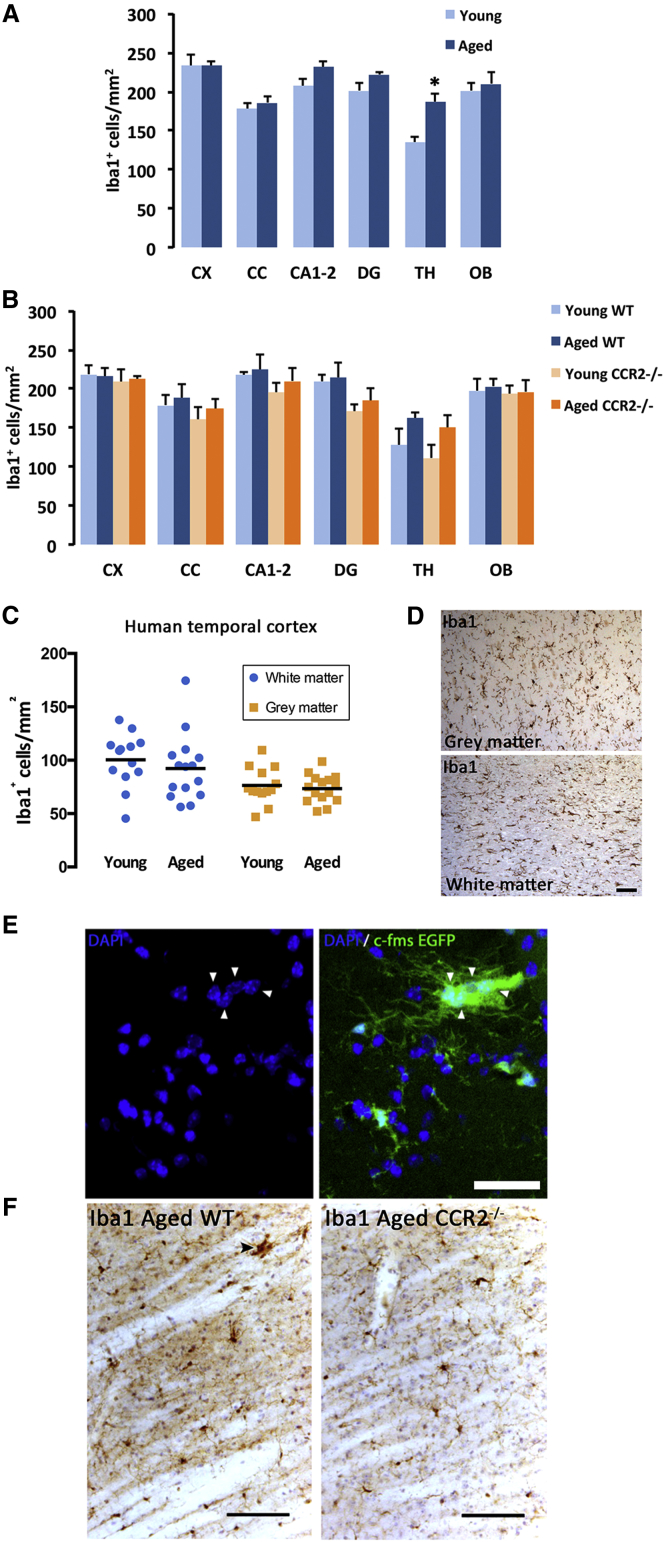
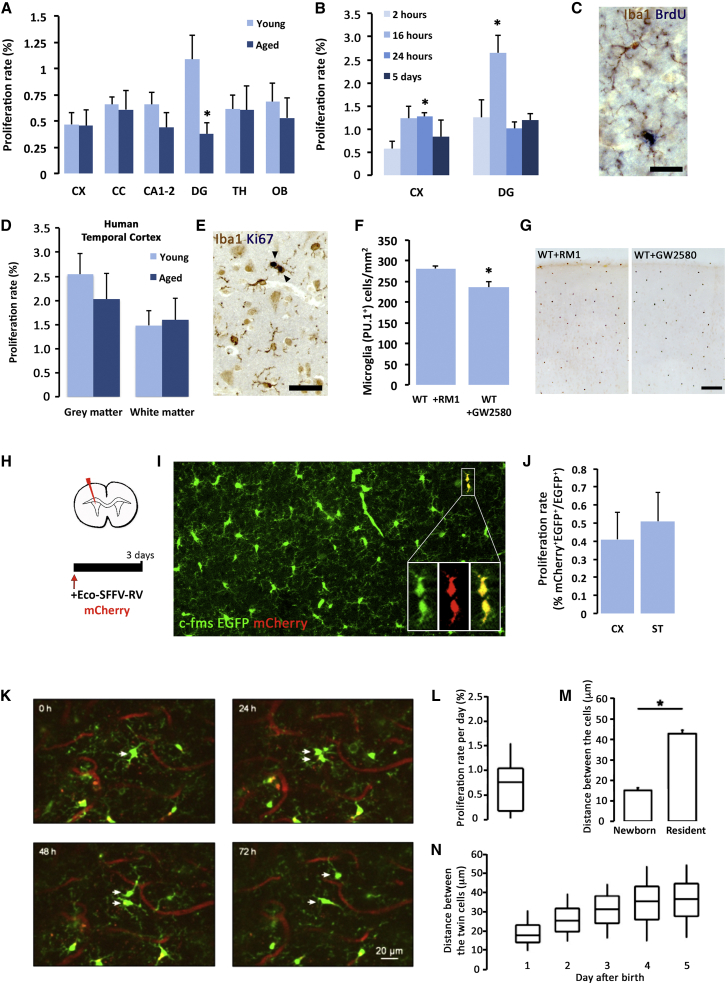
Microglia play key roles in brain development, homeostasis, and function, and it is widely assumed that the adult population is long lived and maintained by self-renewal. However, the precise temporal and spatial dynamics of the microglial population are unknown. We show in mice and humans that the turnover of microglia is remarkably fast, allowing the whole population to be renewed several times during a lifetime. The number of microglial cells remains steady from late postnatal stages until aging and is maintained by the spatial and temporal coupling of proliferation and apoptosis, as shown by pulse-chase studies, chronic in vivo imaging of microglia, and the use of mouse models of dysregulated apoptosis. Our results reveal that the microglial population is constantly and rapidly remodeled, expanding our understanding of its role in the maintenance of brain homeostasis.
Keywords: BrdU; CSF1R; CX3CR1; Macgreen; RNA-seq; Vav-Bcl2; self-renewal.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Alliot F., Godin I., Pessac B. Microglia derive from progenitors, originating from the yolk sac, and which proliferate in the brain. Brain Res. Dev. Brain Res. 1999;117:145–152. - PubMed
-
- Bruttger J., Karram K., Wörtge S., Regen T., Marini F., Hoppmann N., Klein M., Blank T., Yona S., Wolf Y. Genetic cell ablation reveals clusters of local self-renewing microglia in the mammalian central nervous system. Immunity. 2015;43:92–106. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
