

Antigen presenting capacity of murine splenic myeloid cells
- PMID: 28077081
- PMCID: PMC5225582
- DOI: 10.1186/s12865-016-0186-4
Antigen presenting capacity of murine splenic myeloid cells
Abstract
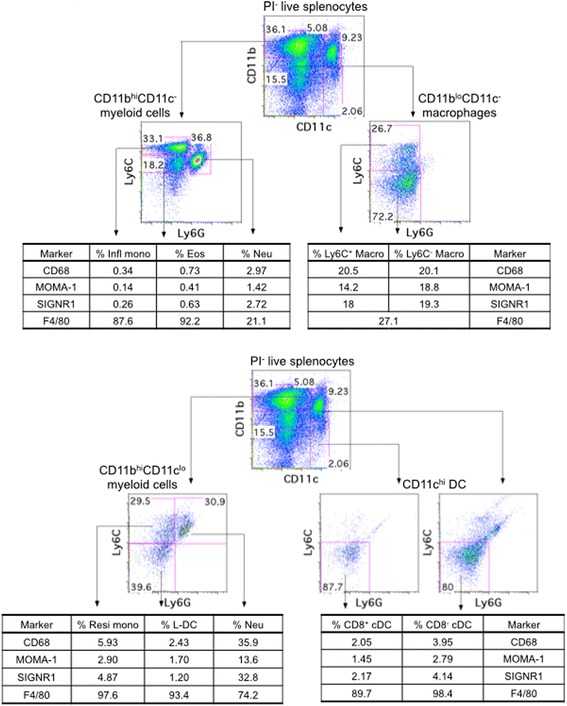
Background: The spleen is an important site for hematopoiesis. It supports development of myeloid cells from bone marrow-derived precursors entering from blood. Myeloid subsets in spleen are not well characterised although dendritic cell (DC) subsets are clearly defined in terms of phenotype, development and functional role. Recently a novel dendritic-like cell type in spleen named 'L-DC' was distinguished from other known dendritic and myeloid cells by its distinct phenotype and developmental origin. That study also redefined splenic eosinophils as well as resident and inflammatory monocytes in spleen.
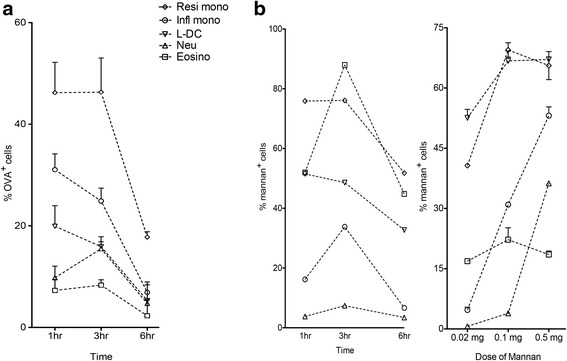
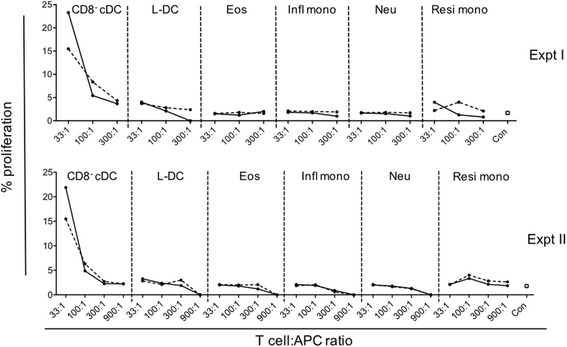
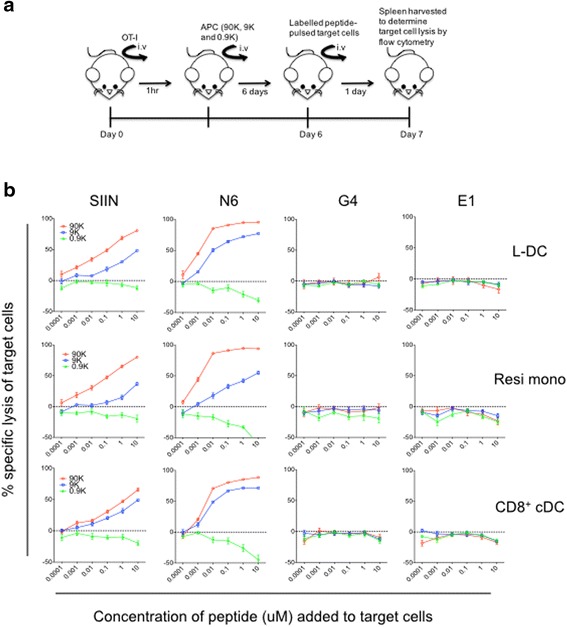
Results: L-DC are shown to be distinct from known splenic macrophages and monocyte subsets. Using a new flow cytometric procedure, it has been possible to identify and isolate L-DC in order to assess their functional competence and ability to activate T cells both in vivo and in vitro. L-DC are readily accessible to antigen given intravenously through receptor-mediated endocytosis. They are also capable of CD8+ T cell activation through antigen cross presentation, with subsequent induction of cytotoxic effector T cells. L-DC are MHCII- cells and unable to activate CD4+ T cells, a property which clearly distinguishes them from conventional DC. The myeloid subsets of resident monocytes, inflammatory monocytes, neutrophils and eosinophils, were found to have varying capacities to take up antigen, but were uniformly unable to activate either CD4+ T cells or CD8+ T cells.
Conclusion: The results presented here demonstrate that L-DC in spleen are distinct from other myeloid cells in that they can process antigen for CD8+ T cell activation and induction of cytotoxic effector function, while both L-DC and myeloid subsets remain unable to activate CD4+ T cells. The L-DC subset in spleen is therefore distinct as an antigen presenting cell.
Keywords: Antigen presentation/processing; Dendritic cells; Myeloid cells; Spleen.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
