

Effector-attenuating Substitutions That Maintain Antibody Stability and Reduce Toxicity in Mice
- PMID: 28077575
- PMCID: PMC5339770
- DOI: 10.1074/jbc.M116.767749
Effector-attenuating Substitutions That Maintain Antibody Stability and Reduce Toxicity in Mice
Abstract
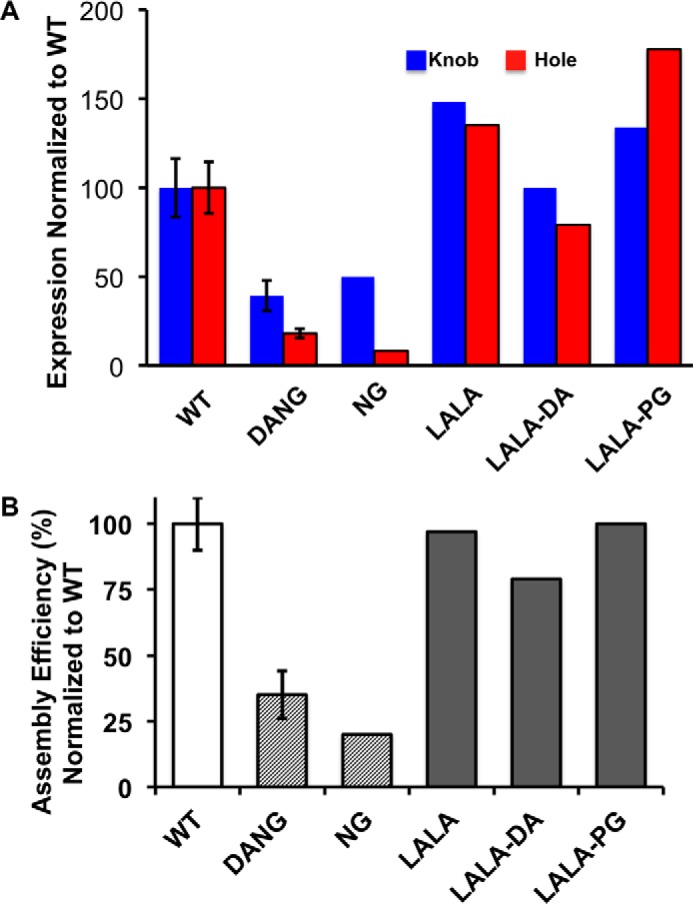
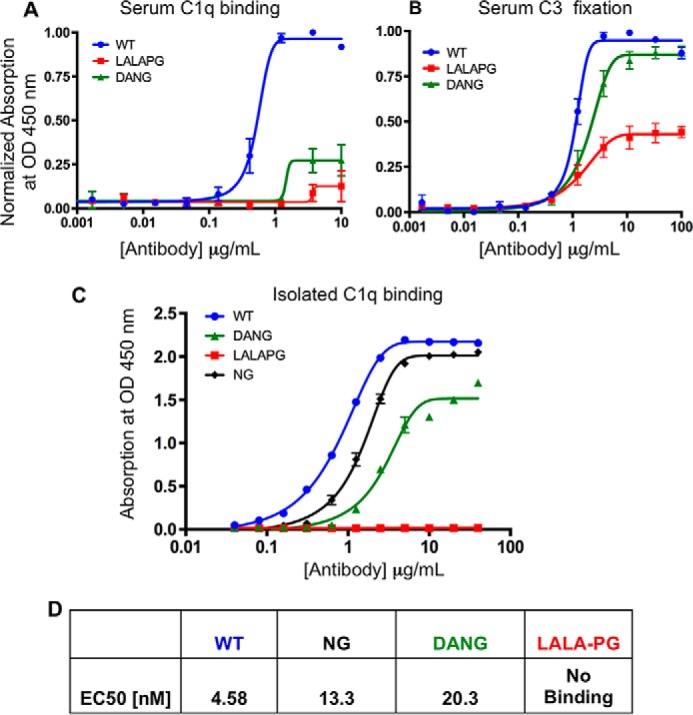
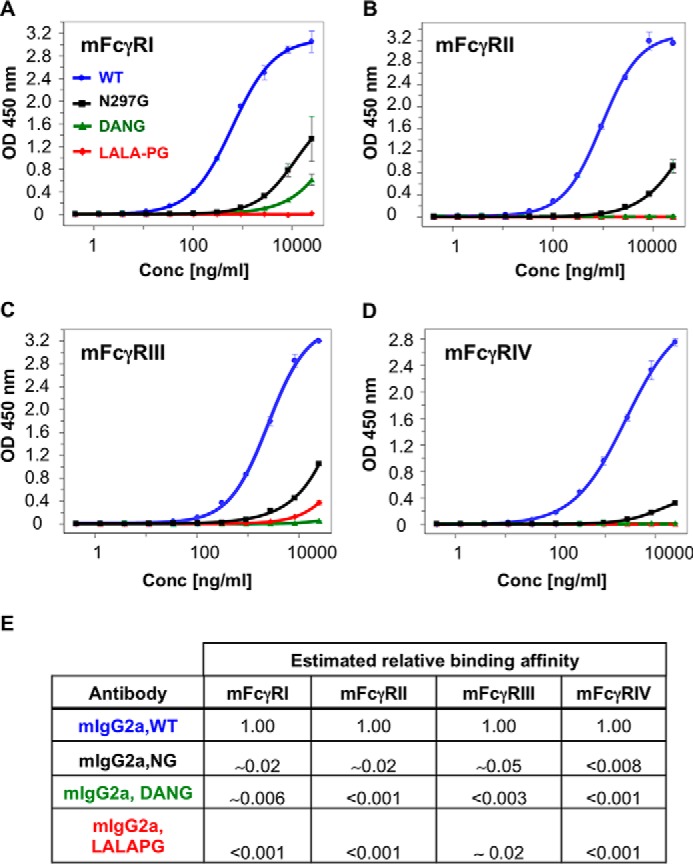
The antibody Fc region regulates antibody cytotoxic activities and serum half-life. In a therapeutic context, however, the cytotoxic effector function of an antibody is often not desirable and can create safety liabilities by activating native host immune defenses against cells expressing the receptor antigens. Several amino acid changes in the Fc region have been reported to silence or reduce the effector function of antibodies. These earlier studies focused primarily on the interaction of human antibodies with human Fc-γ receptors, and it remains largely unknown how such changes to Fc might translate to the context of a murine antibody. We demonstrate that the commonly used N297G (NG) and D265A, N297G (DANG) variants that are efficacious in attenuating effector function in primates retain potent complement activation capacity in mice, leading to safety liabilities in murine studies. In contrast, we found an L234A, L235A, P329G (LALA-PG) variant that eliminates complement binding and fixation as well as Fc-γ-dependent, antibody-dependent, cell-mediated cytotoxity in both murine IgG2a and human IgG1. These LALA-PG substitutions allow a more accurate translation of results generated with an "effectorless" antibody between mice and primates. Further, we show that both human and murine antibodies containing the LALA-PG variant have typical pharmacokinetics in rodents and retain thermostability, enabling efficient knobs-into-holes bispecific antibody production and a robust path to generating highly effector-attenuated bispecific antibodies for preclinical studies.
Keywords: Fc receptor; Fc-γ receptor; aglycosylation; antibody engineering; antibody-dependent, cell-mediated cytotoxicity (ADCC); bispecific antibody; complement; complement-dependent cytotoxicity (CDC); protein stability; recombinant protein expression.
© 2017 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
All authors are present or former paid employees of Genentech Inc.
Figures



References
-
- Walsh G. (2014) Biopharmaceutical benchmarks 2014. Nat. Biotechnol. 32, 992–1000 - PubMed
-
- Spiess C., Zhai Q., and Carter P. J. (2015) Alternative molecular formats and therapeutic applications for bispecific antibodies. Mol. Immunol. 67, 95–106 - PubMed
-
- Sheridan C. (2015) Amgen's bispecific antibody puffs across finish line. Nat. Biotechnol. 33, 219–221 - PubMed
-
- Chan A. C., and Carter P. J. (2010) Therapeutic antibodies for autoimmunity and inflammation. Nat. Rev. Immunol. 10, 301–316 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
