

SUN2 Silencing Impairs CD4 T Cell Proliferation and Alters Sensitivity to HIV-1 Infection Independently of Cyclophilin A
- PMID: 28077629
- PMCID: PMC5331816
- DOI: 10.1128/JVI.02303-16
SUN2 Silencing Impairs CD4 T Cell Proliferation and Alters Sensitivity to HIV-1 Infection Independently of Cyclophilin A
Abstract
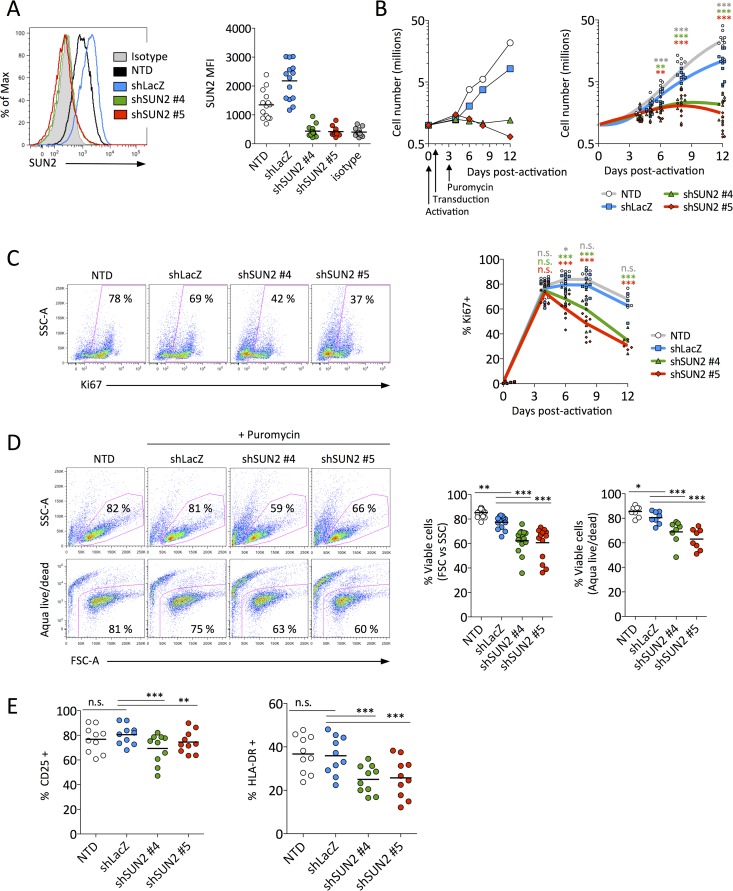
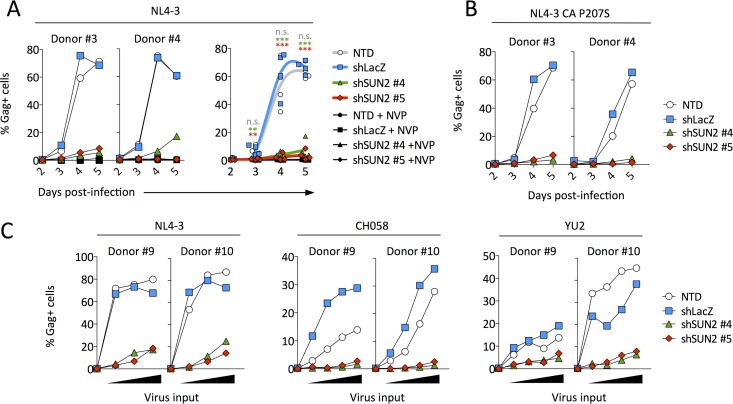
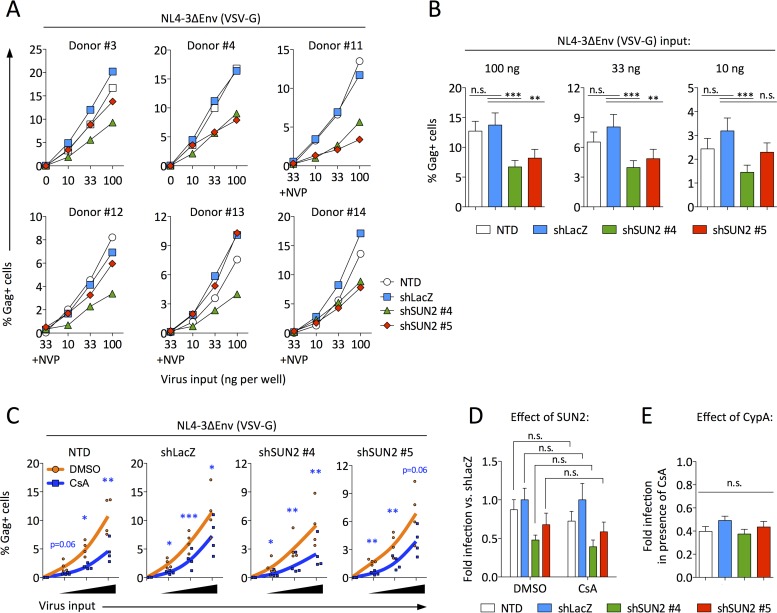
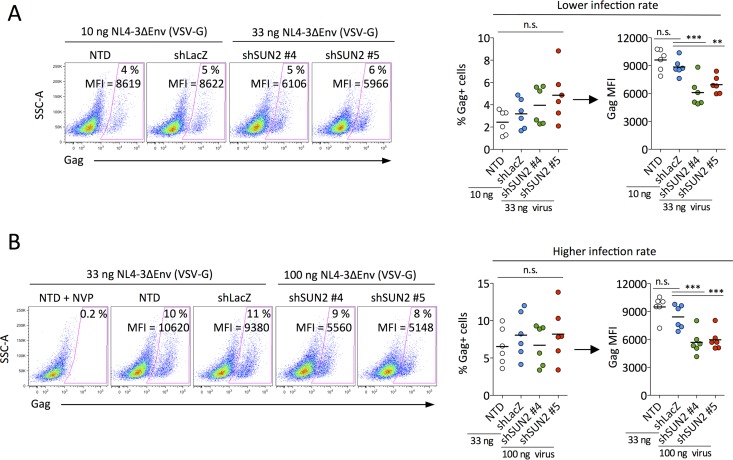
Linker of nucleoskeleton and cytoskeleton (LINC) complexes connect the nucleus to the cytoskeleton in eukaryotic cells. We previously reported that the overexpression of SUN2, an inner nuclear membrane protein and LINC complex component, inhibits HIV infection between the steps of reverse transcription and nuclear import in a capsid-specific manner. We also reported that SUN2 silencing does not modulate HIV infection in several cell lines. Silencing of SUN2 was recently reported to decrease HIV infection of CD4 T cells, an effect which was suggested to result from modulation of cyclophilin A (CypA)-dependent steps of HIV infection. We confirm here that HIV infection of primary CD4 T cells is compromised in the absence of endogenous SUN2, and we extend these findings to additional viral strains. However, we find that CypA is not required for the decreased infection observed in SUN2-silenced cells and, conversely, that endogenous SUN2 is not required for the well-documented positive modulation of HIV infection by CypA. In contrast, CD4 T cells lacking SUN2 exhibit a considerable defect in proliferative capacity and display reduced levels of activation markers and decreased viability. Additionally, SUN2-silenced CD4 T cells that become infected support reduced levels of viral protein expression. Our results demonstrate that SUN2 is required for the optimal activation and proliferation of primary CD4 T cells and suggest that the disruption of these processes explains the contribution of endogenous SUN2 to HIV infection in primary lymphocytes.IMPORTANCE Linker of nucleoskeleton and cytoskeleton (LINC) complexes connect the nucleus to the cytoskeleton. We previously reported that the overexpression of the LINC complex protein SUN2 inhibits HIV infection by targeting the viral capsid and blocking infection before the virus enters the nucleus. A recent report showed that the depletion of endogenous SUN2 in primary CD4 T cells results in decreased HIV infection and that this involves cyclophilin A (CypA), a host protein that interacts with the capsid of HIV to promote infection. We confirm that HIV infection is reduced in CD4 T cells lacking SUN2, but we find no role for CypA. Instead, SUN2 silencing results in CD4 T cells with decreased viability and much lower proliferation rates. Our results show that SUN2 is required for optimal CD4 T cell activation and proliferation and explain the reduced level of HIV infection in the absence of SUN2.
Keywords: CD4 T cell; SUN2; human immunodeficiency virus.
Copyright © 2017 American Society for Microbiology.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
