

Symbiont-induced odorant binding proteins mediate insect host hematopoiesis
- PMID: 28079523
- PMCID: PMC5231409
- DOI: 10.7554/eLife.19535
Symbiont-induced odorant binding proteins mediate insect host hematopoiesis
Abstract
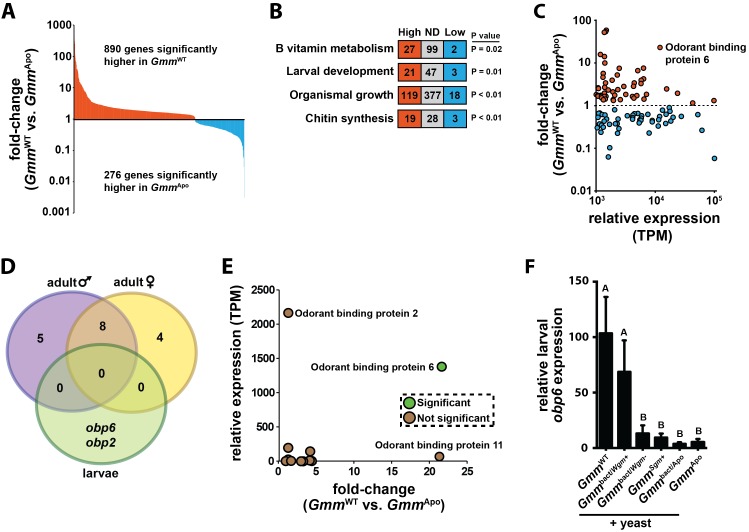
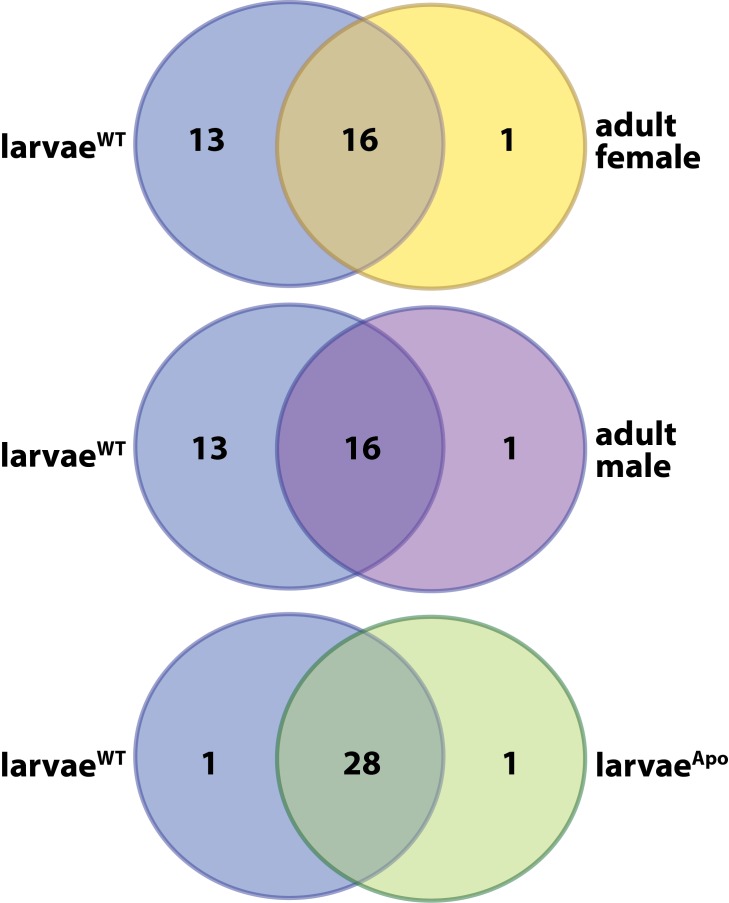
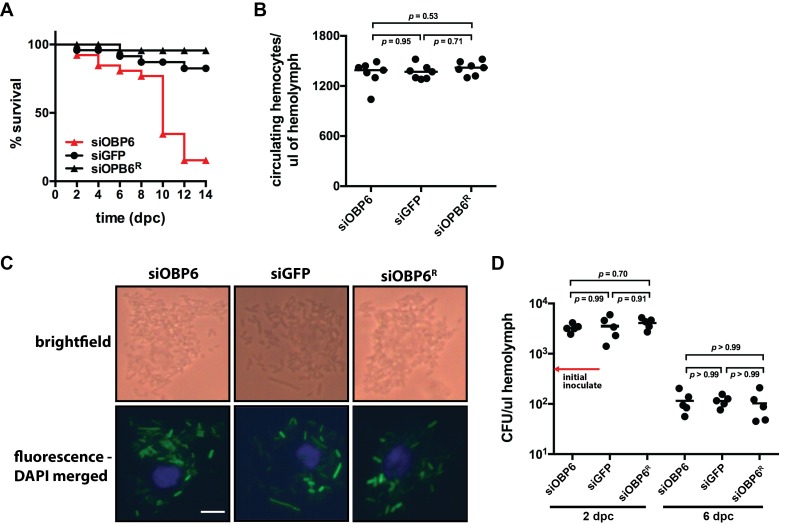
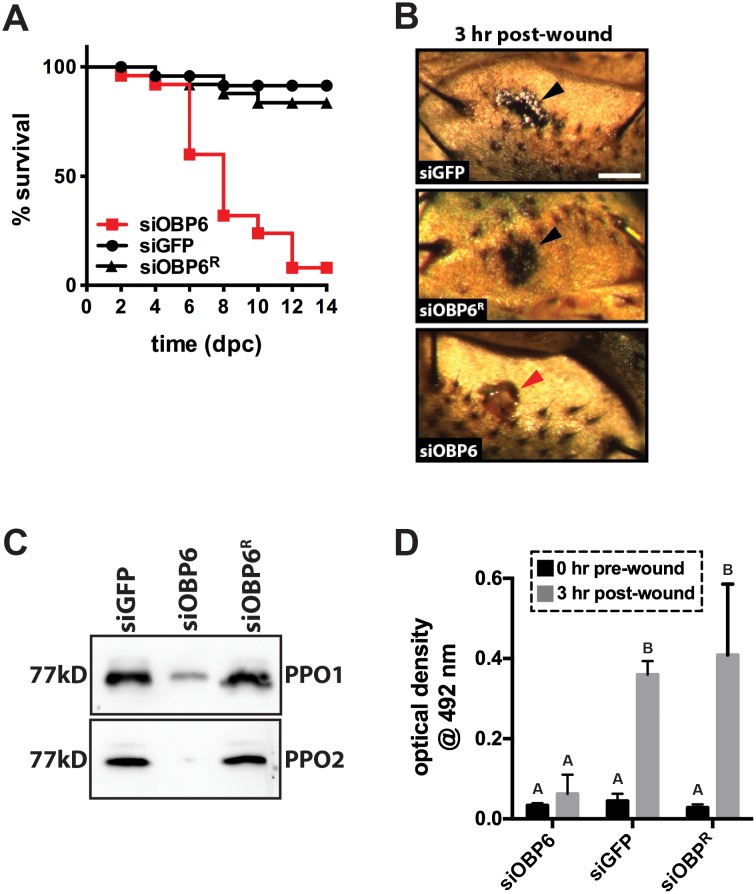
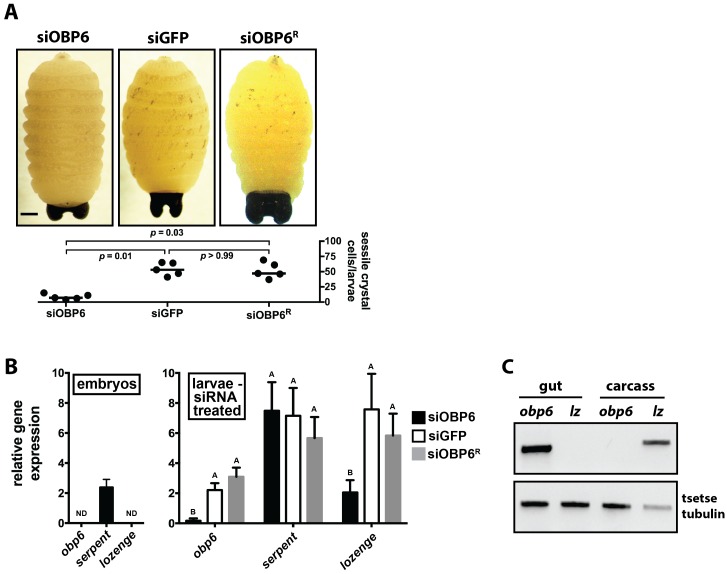
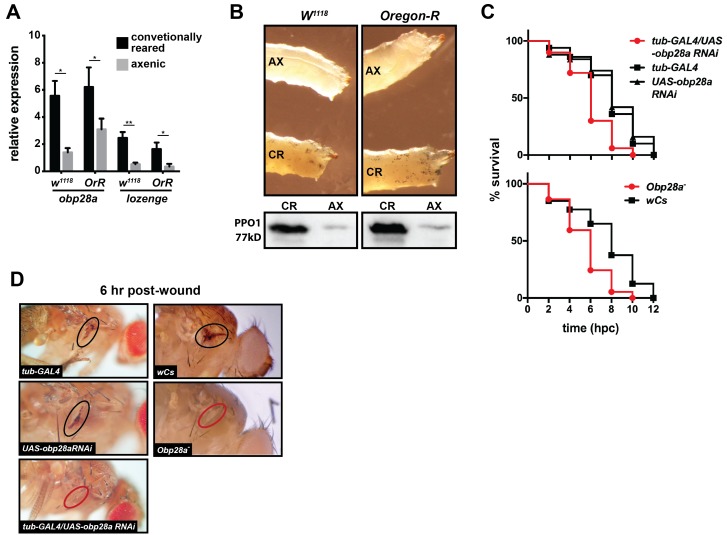
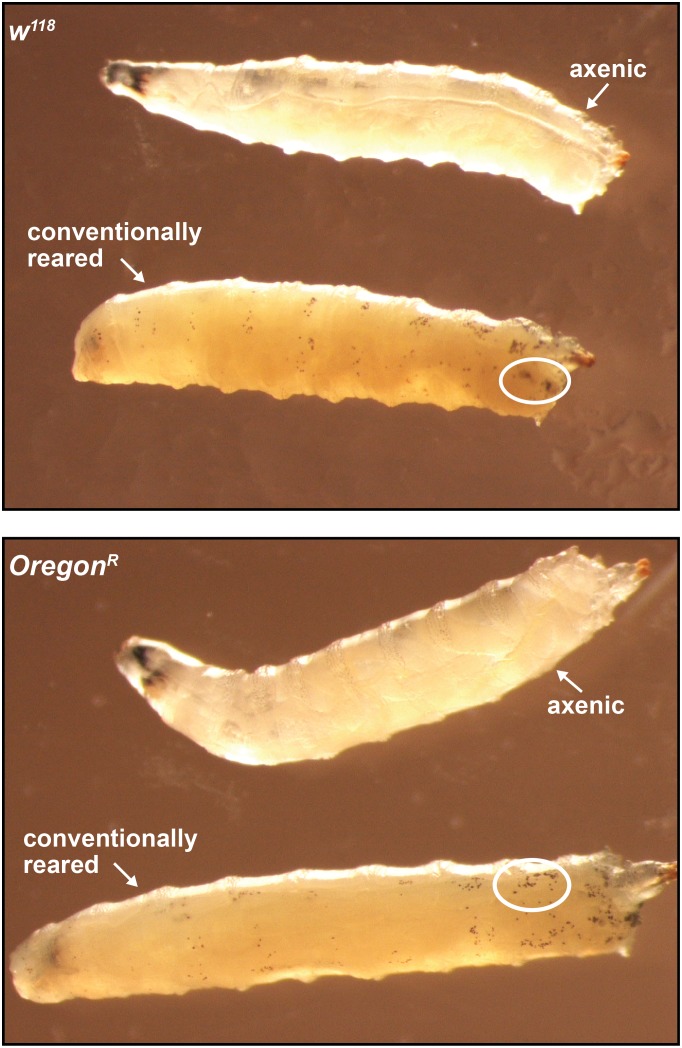
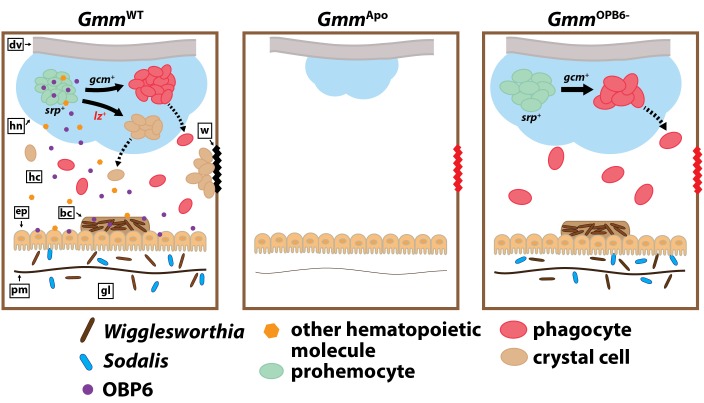
Symbiotic bacteria assist in maintaining homeostasis of the animal immune system. However, the molecular mechanisms that underlie symbiont-mediated host immunity are largely unknown. Tsetse flies (Glossina spp.) house maternally transmitted symbionts that regulate the development and function of their host's immune system. Herein we demonstrate that the obligate mutualist, Wigglesworthia, up-regulates expression of odorant binding protein six in the gut of intrauterine tsetse larvae. This process is necessary and sufficient to induce systemic expression of the hematopoietic RUNX transcription factor lozenge and the subsequent production of crystal cells, which actuate the melanotic immune response in adult tsetse. Larval Drosophila's indigenous microbiota, which is acquired from the environment, regulates an orthologous hematopoietic pathway in their host. These findings provide insight into the molecular mechanisms that underlie enteric symbiont-stimulated systemic immune system development, and indicate that these processes are evolutionarily conserved despite the divergent nature of host-symbiont interactions in these model systems.
Keywords: D. melanogaster; crystal cell; developmental biology; hematopoiesis; immunology; odorant binding protein; stem cells; symbiont; tsetse fly.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
Comment in
-
Protection from within.Elife. 2017 Jan 12;6:e24111. doi: 10.7554/eLife.24111. Elife. 2017. PMID: 28079522 Free PMC article.
References
-
- Alam U, Medlock J, Brelsfoard C, Pais R, Lohs C, Balmand S, Carnogursky J, Heddi A, Takac P, Galvani A, Aksoy S. Wolbachia symbiont infections induce strong cytoplasmic incompatibility in the tsetse fly Glossina morsitans. PLoS Pathogens. 2011;7:e1002415. doi: 10.1371/journal.ppat.1002415. - DOI - PMC - PubMed
-
- Attardo GM, Benoit JB, Michalkova V, Patrick KR, Krause TB, Aksoy S. The homeodomain protein ladybird late regulates synthesis of milk proteins during pregnancy in the tsetse fly (Glossina morsitans) PLoS Neglected Tropical Diseases. 2014;8:e2645. doi: 10.1371/journal.pntd.0002645. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
