

Maternal Supply of Cas9 to Zygotes Facilitates the Efficient Generation of Site-Specific Mutant Mouse Models
- PMID: 28081254
- PMCID: PMC5231326
- DOI: 10.1371/journal.pone.0169887
Maternal Supply of Cas9 to Zygotes Facilitates the Efficient Generation of Site-Specific Mutant Mouse Models
Abstract
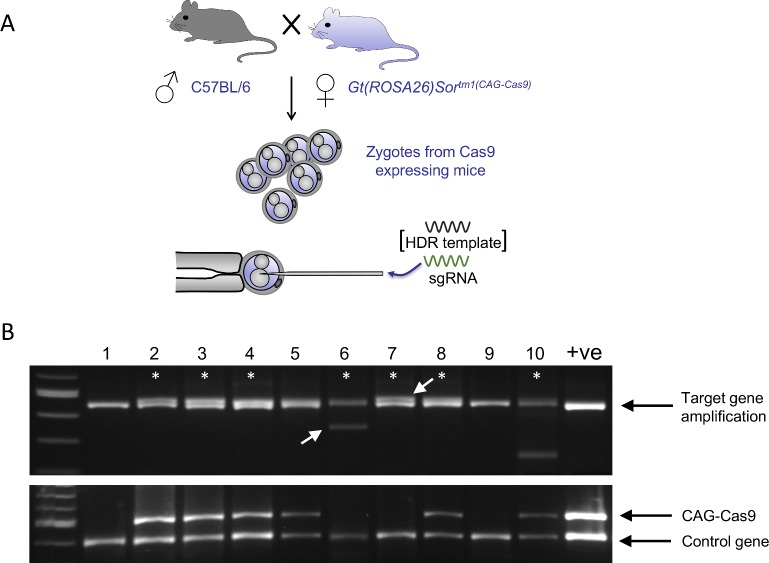
Genome manipulation in the mouse via microinjection of CRISPR/Cas9 site-specific nucleases has allowed the production time for genetically modified mouse models to be significantly reduced. Successful genome manipulation in the mouse has already been reported using Cas9 supplied by microinjection of a DNA construct, in vitro transcribed mRNA and recombinant protein. Recently the use of transgenic strains of mice overexpressing Cas9 has been shown to facilitate site-specific mutagenesis via maternal supply to zygotes and this route may provide an alternative to exogenous supply. We have investigated the feasibility of supplying Cas9 genetically in more detail and for this purpose we report the generation of a transgenic mice which overexpress Cas9 ubiquitously, via a CAG-Cas9 transgene targeted to the Gt(ROSA26)Sor locus. We show that zygotes prepared from female mice harbouring this transgene are sufficiently loaded with maternally contributed Cas9 for efficient production of embryos and mice harbouring indel, genomic deletion and knock-in alleles by microinjection of guide RNAs and templates alone. We compare the mutagenesis rates and efficacy of mutagenesis using this genetic supply with exogenous Cas9 supply by either mRNA or protein microinjection. In general, we report increased generation rates of knock-in alleles and show that the levels of mutagenesis at certain genome target sites are significantly higher and more consistent when Cas9 is supplied genetically relative to exogenous supply.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
