

Hepatocyte-specific PPARA expression exclusively promotes agonist-induced cell proliferation without influence from nonparenchymal cells
- PMID: 28082284
- PMCID: PMC5401987
- DOI: 10.1152/ajpgi.00205.2016
Hepatocyte-specific PPARA expression exclusively promotes agonist-induced cell proliferation without influence from nonparenchymal cells
Abstract
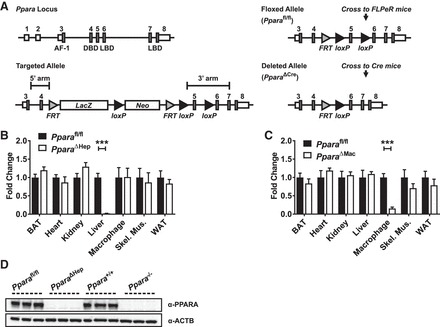
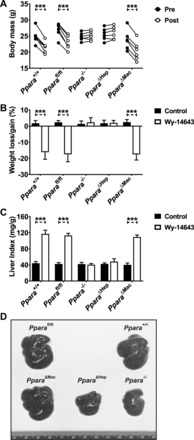
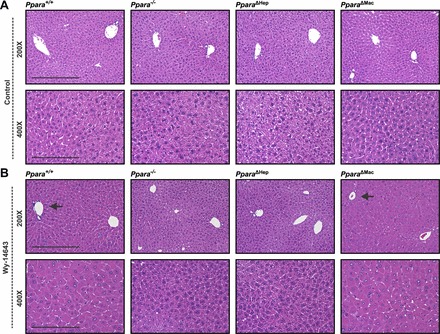
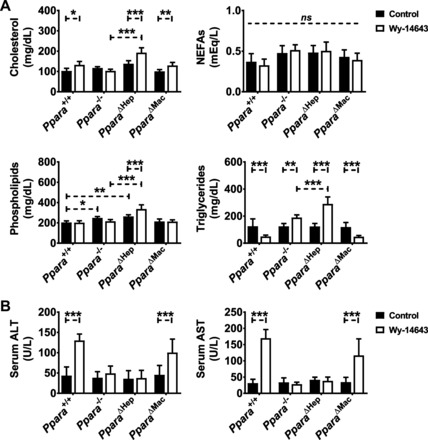
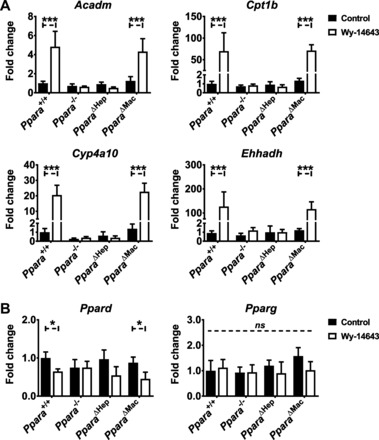
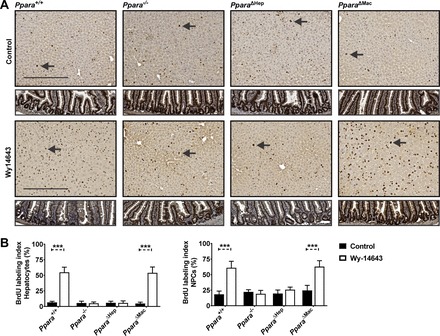
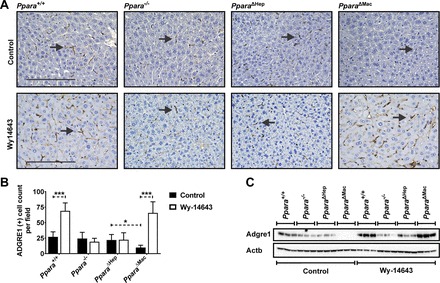
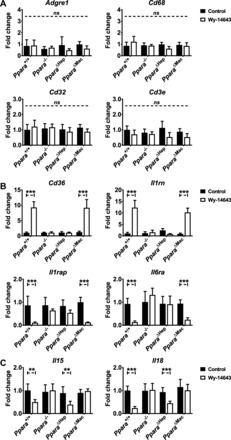
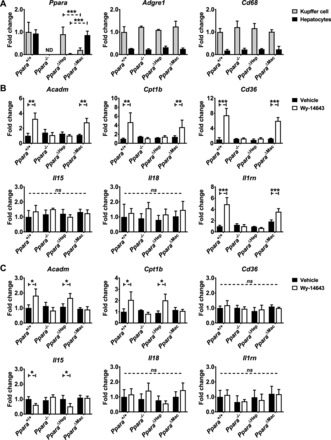
Peroxisome proliferator-activated receptor-α (PPARA) is a nuclear transcription factor and key mediator of systemic lipid metabolism. Prolonged activation in rodents causes hepatocyte proliferation and hepatocellular carcinoma. Little is known about the contribution of nonparenchymal cells (NPCs) to PPARA-mediated cell proliferation. NPC contribution to PPARA agonist-induced hepatomegaly was assessed in hepatocyte (Ppara△Hep)- and macrophage (Ppara△Mac)-specific Ppara null mice. Mice were treated with the agonist Wy-14643 for 14 days, and response of conditional null mice was compared with conventional knockout mice (Ppara-/- ). Wy-14643 treatment caused weight loss and severe hepatomegaly in wild-type and Ppara△Mac mice, and histological analysis revealed characteristic hepatocyte swelling; Ppara△Hep and Ppara-/- mice were protected from these effects. Ppara△Mac serum chemistries, as well as aspartate aminotransferase and alanine aminotransferase levels, matched wild-type mice. Agonist-treated Ppara△Hep mice had elevated serum cholesterol, phospholipids, and triglycerides when compared with Ppara-/- mice, indicating a possible role for extrahepatic PPARA in regulating circulating lipid levels. BrdU labeling confirmed increased cell proliferation only in wild-type and Ppara△Mac mice. Macrophage PPARA disruption did not impact agonist-induced upregulation of lipid metabolism, cell proliferation, or DNA damage and repair-related gene expression, whereas gene expression was repressed in Ppara△Hep mice. Interestingly, downregulation of inflammatory cytokines IL-15 and IL-18 was dependent on macrophage PPARA. Cell type-specific regulation of target genes was confirmed in primary hepatocytes and Kupffer cells. These studies conclusively show that cell proliferation is mediated exclusively by PPARA activation in hepatocytes and that Kupffer cell PPARA has an important role in mediating the anti-inflammatory effects of PPARA agonists.
Keywords: Kupffer cell; Wy-14643; fibrate; peroxisome proliferator-activated receptor-α; proliferation.
Copyright © 2017 the American Physiological Society.
Figures

References
-
- Bustin SA, Benes V, Garson JA, Hellemans J, Huggett J, Kubista M, Mueller R, Nolan T, Pfaffl MW, Shipley GL, Vandesompele J, Wittwer CT. The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin Chem 55: 611–622, 2009. doi: 10.1373/clinchem.2008.112797. - DOI - PubMed
-
- Desvergne B, Wahli W. Peroxisome proliferator-activated receptors: nuclear control of metabolism. Endocr Rev 20: 649–688, 1999. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous