

Phylogenomics and Plastome Evolution of Tropical Forest Grasses (Leptaspis, Streptochaeta: Poaceae)
- PMID: 28083012
- PMCID: PMC5186769
- DOI: 10.3389/fpls.2016.01993
Phylogenomics and Plastome Evolution of Tropical Forest Grasses (Leptaspis, Streptochaeta: Poaceae)
Abstract
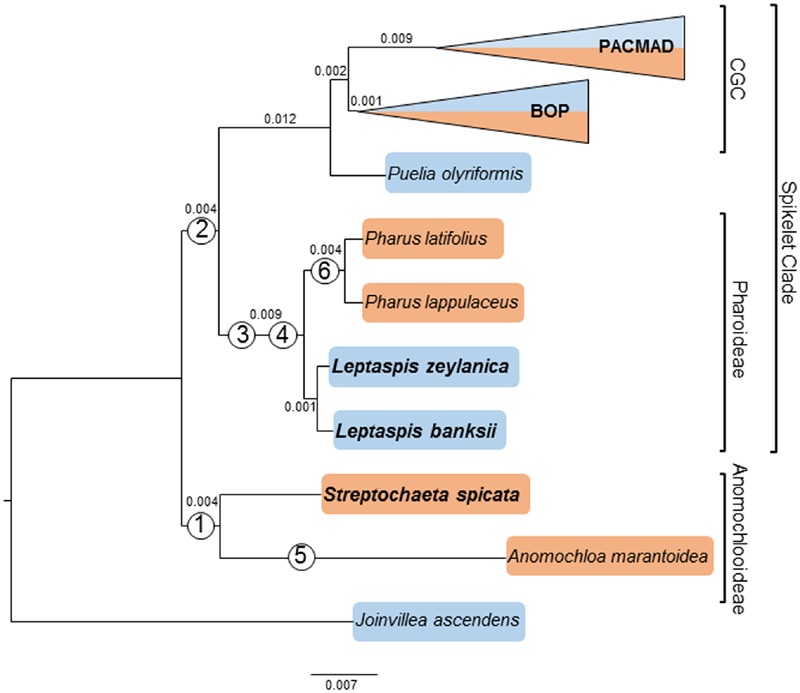
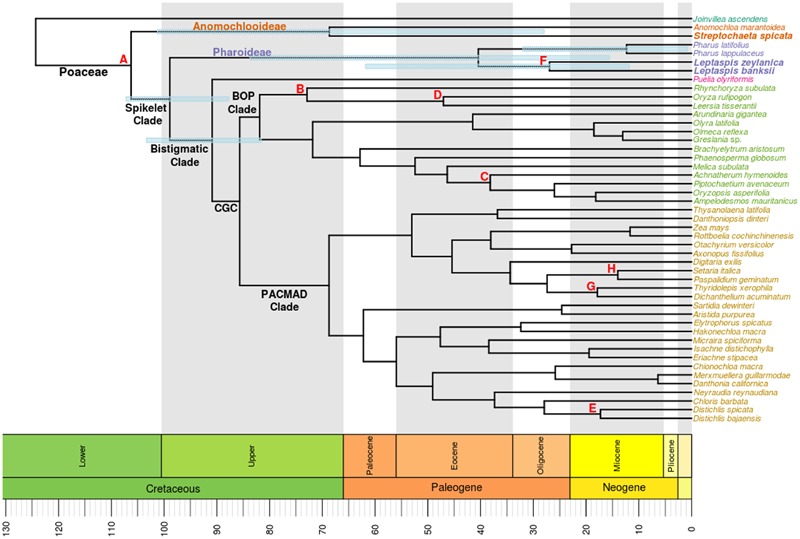
Studies of complete plastomes have proven informative for our understanding of the molecular evolution and phylogenomics of grasses. In this study, a plastome phylogenomic analysis sampled species from lineages of deeply diverging grasses including Streptochaeta spicata (Anomochlooideae), Leptaspis banksii, and L. zeylanica (both Pharoideae). Plastomes from next generation sequences for three species were assembled by de novo methods. The unambiguously aligned coding and non-coding sequences of the entire plastomes were aligned with those from 43 other grasses and the outgroup Joinvillea ascendens. Outgroup sampling of grasses has previously posed a challenge for plastome phylogenomic studies because of major rearrangements of the plastome. Here, over 81,000 bases of homologous sequence were aligned for phylogenomic and divergence estimation analyses. Rare genomic changes, including persistently long ψycf1 and ψycf2 loci, the loss of the rpoC1 intron, and a 21 base tandem repeat insert in the coding sequence for rps19 defined branch points in the grass phylogeny. Marked differences were seen in the topologies inferred from the complete plastome and two gene matrices, and mean maximum likelihood support values for the former were 10% higher. In the full plastome phylogenomic analyses, the two species of Anomochlooideae were monophyletic. Leptaspis and Pharus were found to be reciprocally monophyletic, with the estimated divergence of two Leptaspis species preceding those of Pharus by over 14 Ma, consistent with historical biogeography. Our estimates for deep divergences among grasses were older than previous such estimates, likely influenced by more complete taxonomic and molecular sampling and the use of recently available or previously unused fossil calibration points.
Keywords: Anomochlooideae; Pharoideae; Poaceae; plastid genome; plastome phylogenomics.
Figures
References
-
- Akaike H. (1974). A new look at the statistical model identification. IEEE Trans. Autom. Control 19 716–723. 10.1109/TAC.1974.1100705 - DOI
-
- Bell Mark A., Lloyd G. T. (2014). strap: Stratigraphic Tree Analysis for Palaeontology. R package version 1.4. Available at: http://CRAN.R-project.org/package=strap
-
- Bouchenak-Khelladi Y., Muasya A. M., Linder H. P. (2014). A revised evolutionary history of Poales: origins and diversification. Bot. J. Linn. Soc. 175 4–16. 10.1111/boj.12160 - DOI
-
- Bouchenak-Khelladi Y., Verboom G. A., Savolainen V., Hodkinson T. (2010). Biogeography of the grasses (Poaceae): a phylogenetic approach to reveal evolutionary history in geographical space and geological time. Bot. J. Linn. Soc. 162 543–557. 10.1111/j.1095-8339.2010.01041.x - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous