

Mitochondrial dynamics as regulators of cancer biology
- PMID: 28083595
- PMCID: PMC5419868
- DOI: 10.1007/s00018-016-2451-3
Mitochondrial dynamics as regulators of cancer biology
Abstract
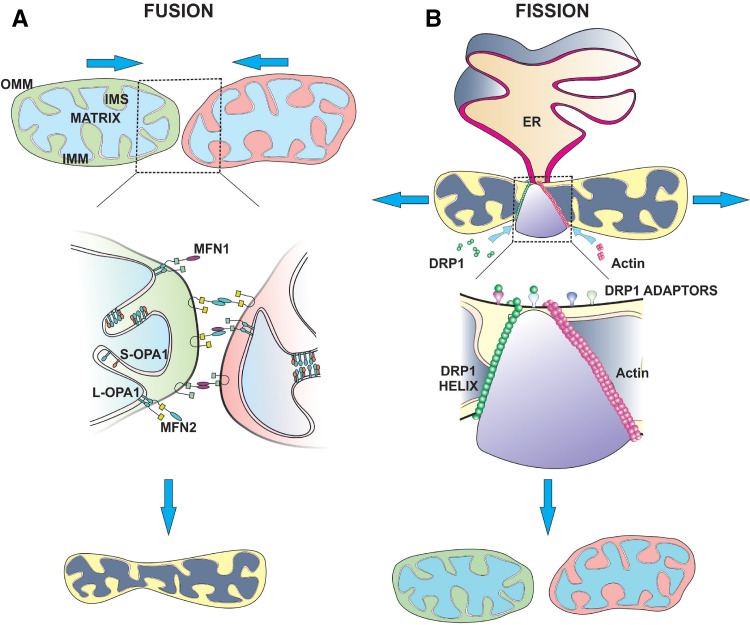
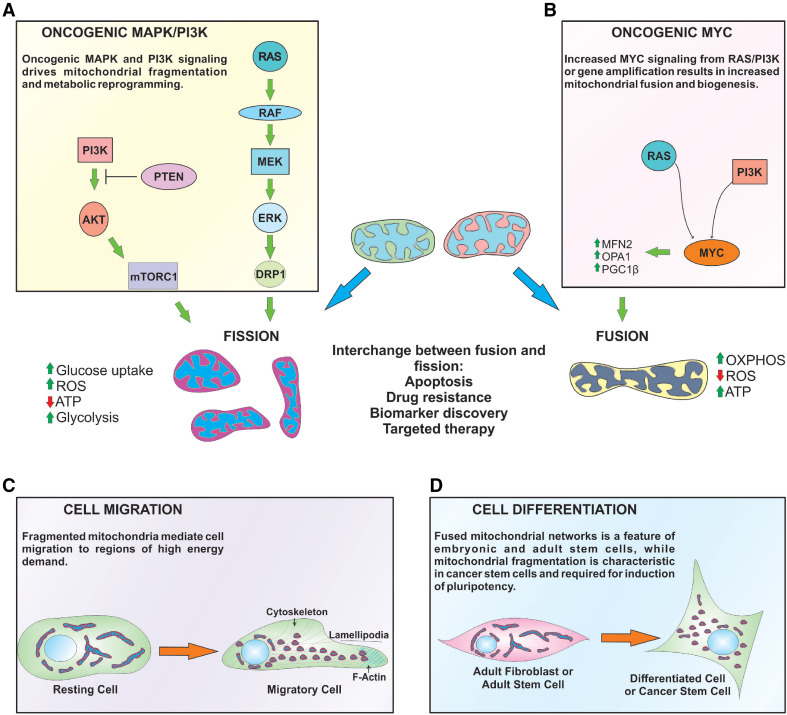
Mitochondria are dynamic organelles that supply energy required to drive key cellular processes, such as survival, proliferation, and migration. Critical to all of these processes are changes in mitochondrial architecture, a mechanical mechanism encompassing both fusion and fragmentation (fission) of the mitochondrial network. Changes to mitochondrial shape, size, and localization occur in a regulated manner to maintain energy and metabolic homeostasis, while deregulation of mitochondrial dynamics is associated with the onset of metabolic dysfunction and disease. In cancers, oncogenic signals that drive excessive proliferation, increase intracellular stress, and limit nutrient supply are all able to alter the bioenergetic and biosynthetic requirements of cancer cells. Consequently, mitochondrial function and shape rapidly adapt to these hostile conditions to support cancer cell proliferation and evade activation of cell death programs. In this review, we will discuss the molecular mechanisms governing mitochondrial dynamics and integrate recent insights into how changes in mitochondrial shape affect cellular migration, differentiation, apoptosis, and opportunities for the development of novel targeted cancer therapies.
Keywords: Apoptosis; Cancer; Differentiation; Migration; Mitochondrial dynamics; Oncogenic signaling.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
