

Proteoliposomal formulations of an HIV-1 gp41-based miniprotein elicit a lipid-dependent immunodominant response overlapping the 2F5 binding motif
- PMID: 28084464
- PMCID: PMC5234007
- DOI: 10.1038/srep40800
Proteoliposomal formulations of an HIV-1 gp41-based miniprotein elicit a lipid-dependent immunodominant response overlapping the 2F5 binding motif
Abstract
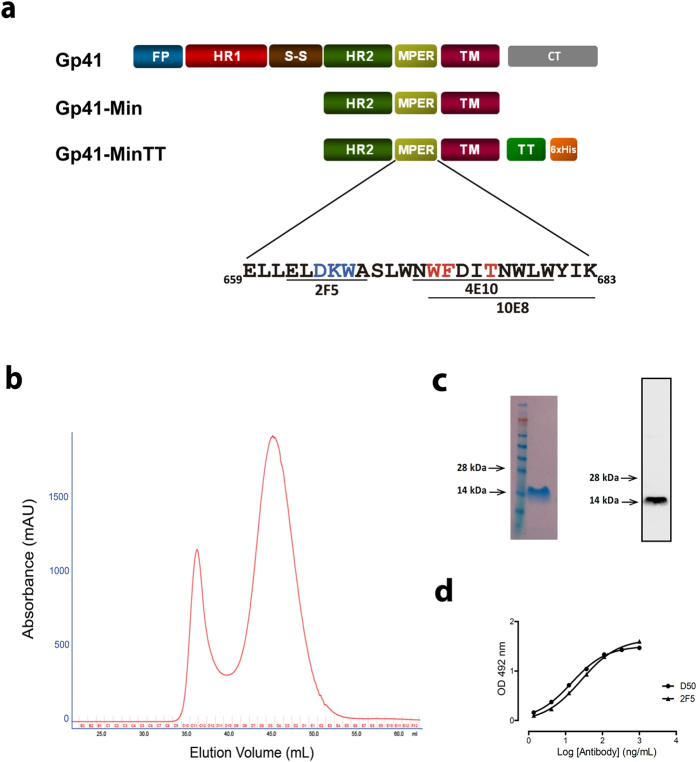
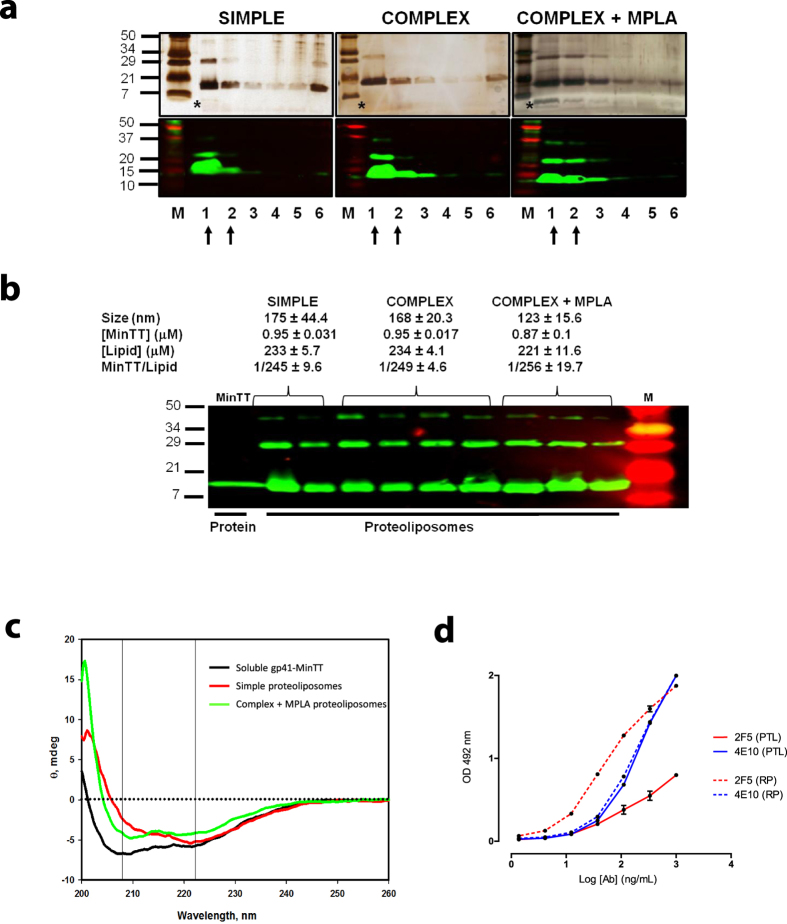
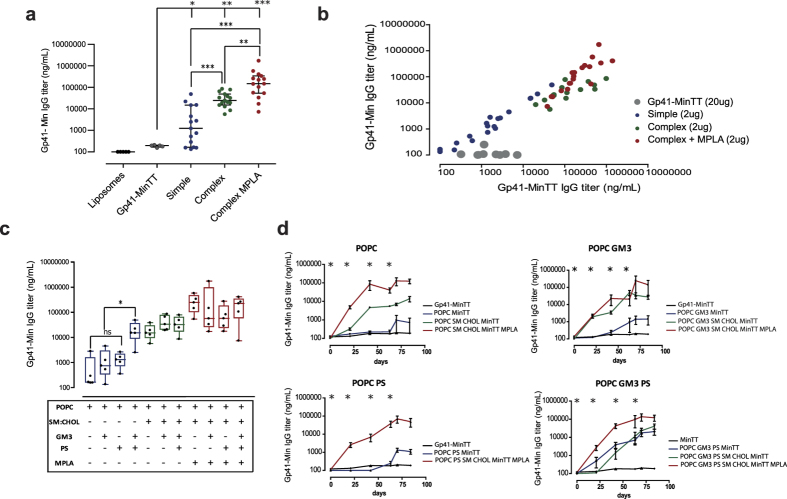
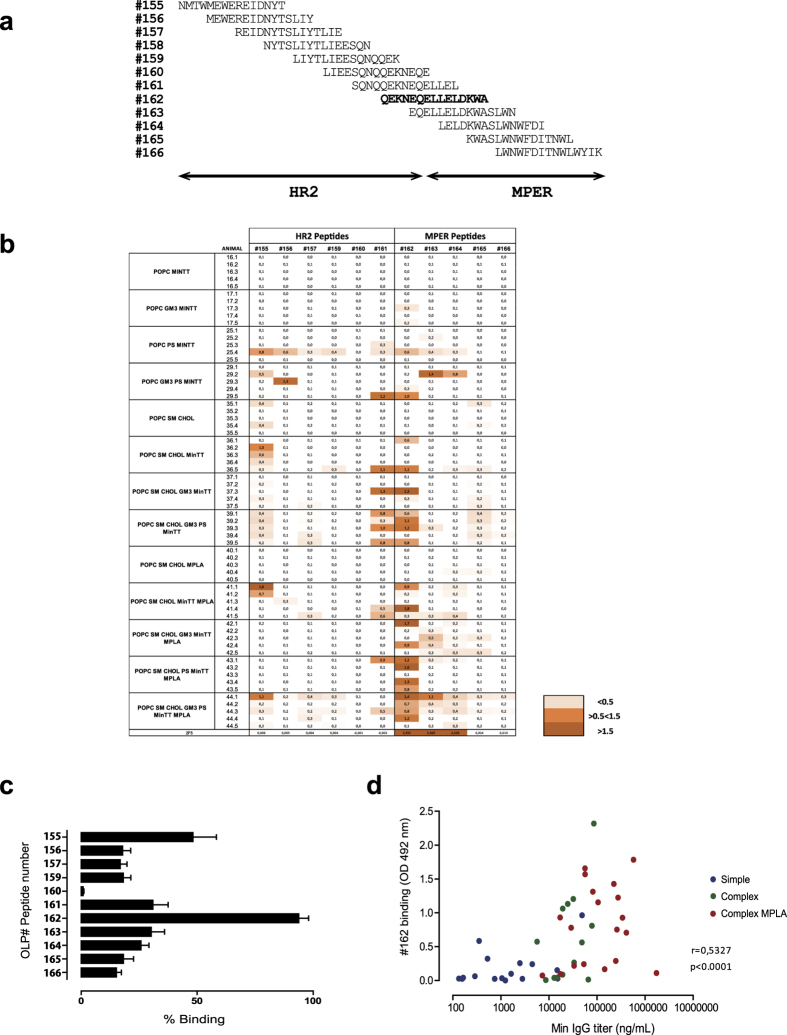
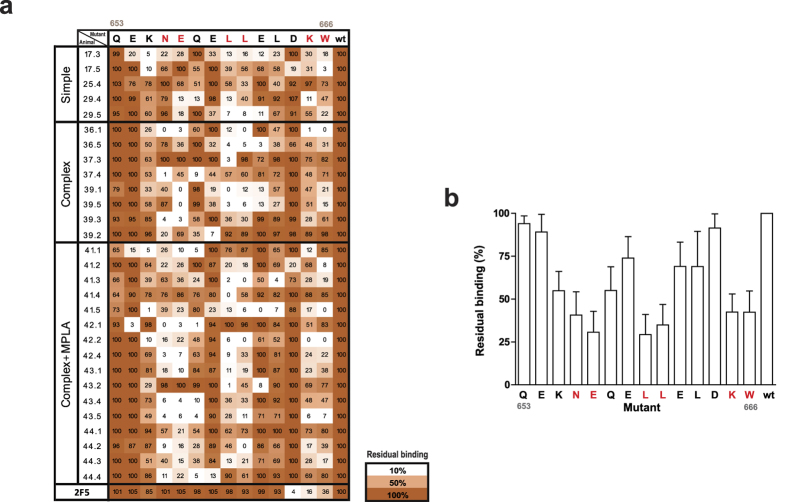
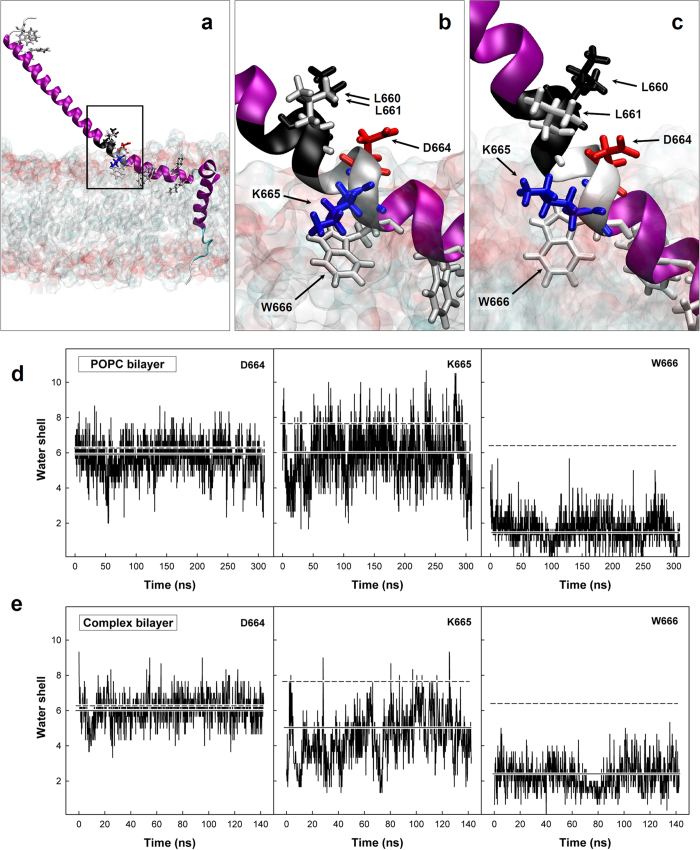
The HIV-1 gp41 Membrane Proximal External Region (MPER) is recognized by broadly neutralizing antibodies and represents a promising vaccine target. However, MPER immunogenicity and antibody activity are influenced by membrane lipids. To evaluate lipid modulation of MPER immunogenicity, we generated a 1-Palmitoyl-2-oleoylphosphatidylcholine (POPC)-based proteoliposome collection containing combinations of phosphatidylserine (PS), GM3 ganglioside, cholesterol (CHOL), sphingomyelin (SM) and the TLR4 agonist monophosphoryl lipid A (MPLA). A recombinant gp41-derived miniprotein (gp41-MinTT) exposing the MPER and a tetanus toxoid (TT) peptide that favors MHC-II presentation, was successfully incorporated into lipid mixtures (>85%). Immunization of mice with soluble gp41-MinTT exclusively induced responses against the TT peptide, while POPC proteoliposomes generated potent anti-gp41 IgG responses using lower protein doses. The combined addition of PS and GM3 or CHOL/SM to POPC liposomes greatly increased gp41 immunogenicity, which was further enhanced by the addition of MPLA. Responses generated by all proteoliposomes targeted the N-terminal moiety of MPER overlapping the 2F5 neutralizing epitope. Our data show that lipids impact both, the epitope targeted and the magnitude of the response to membrane-dependent antigens, helping to improve MPER-based lipid carriers. Moreover, the identification of immunodominant epitopes allows for the redesign of immunogens targeting MPER neutralizing determinants.
Figures
References
-
- Binley J. M. et al. Comprehensive Cross-Clade Neutralization Analysis of a Panel of Anti-Human Immunodeficiency Virus Type 1 Monoclonal Antibodies Comprehensive Cross-Clade Neutralization Analysis of a Panel of Anti-Human Immunodeficiency Virus Type 1 Monoclonal Antibodies. J. Virol. 78, 13232–13252 (2004). - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
