

The BCL-2 pro-survival protein A1 is dispensable for T cell homeostasis on viral infection
- PMID: 28085151
- PMCID: PMC5344212
- DOI: 10.1038/cdd.2016.155
The BCL-2 pro-survival protein A1 is dispensable for T cell homeostasis on viral infection
Abstract
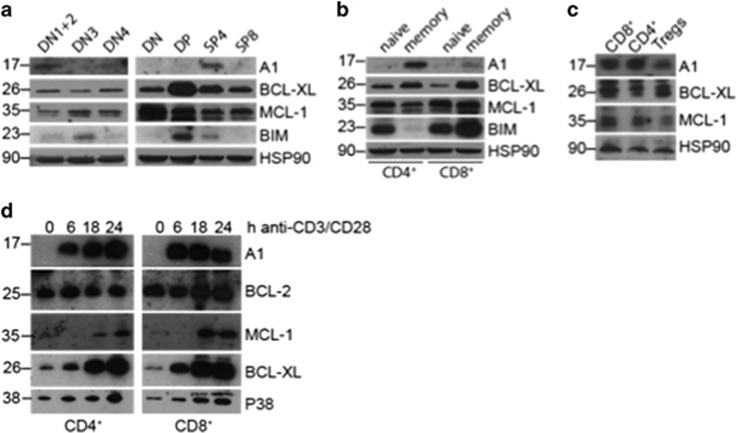
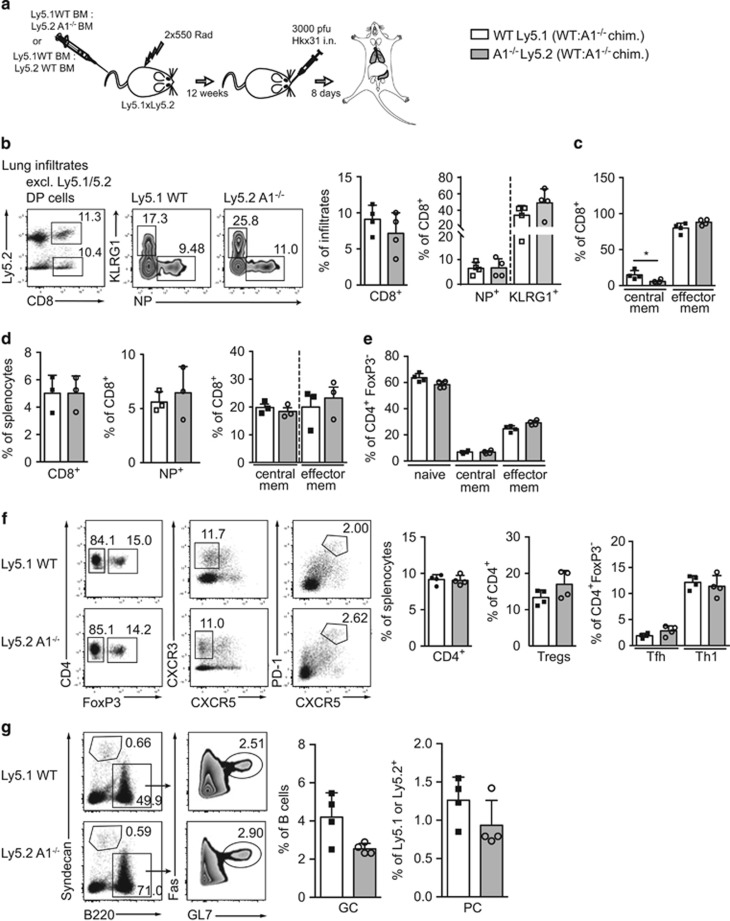
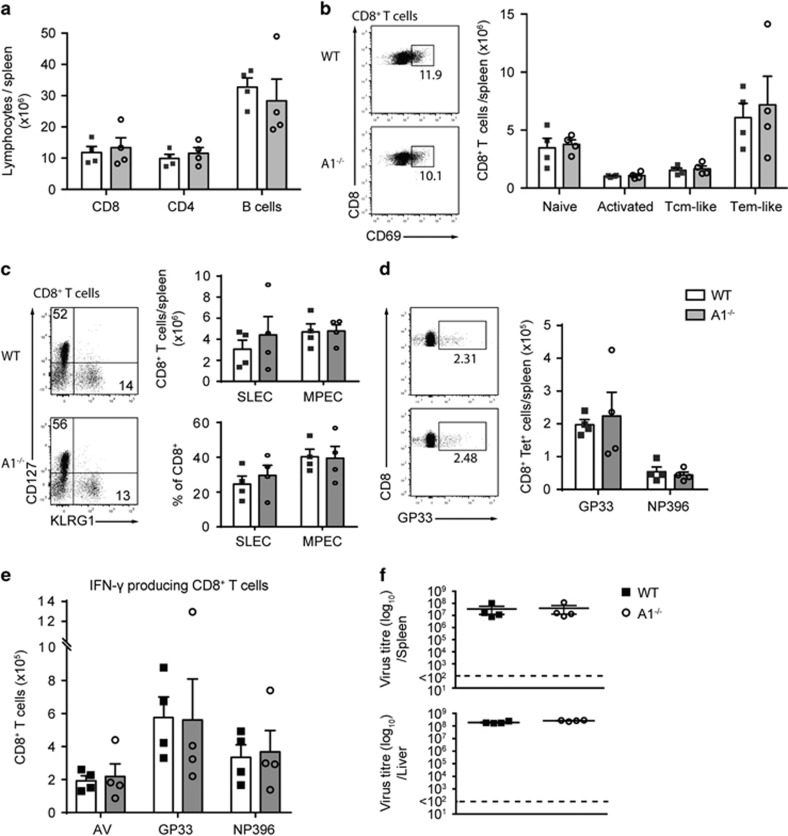
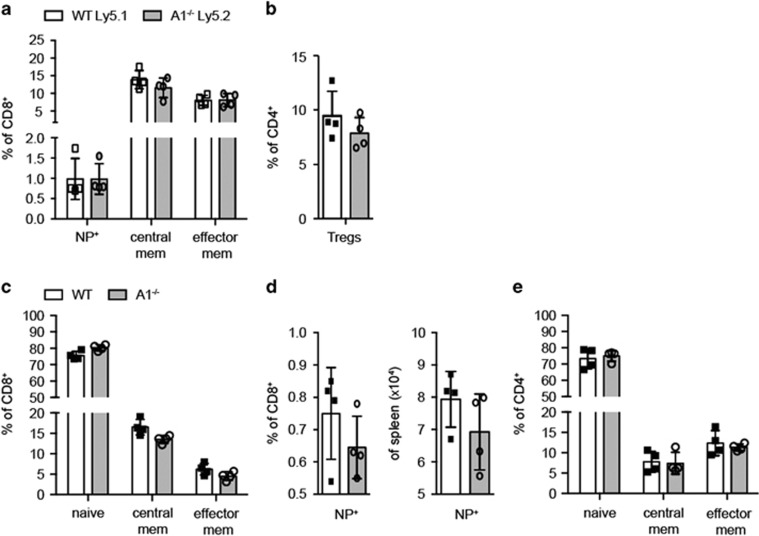
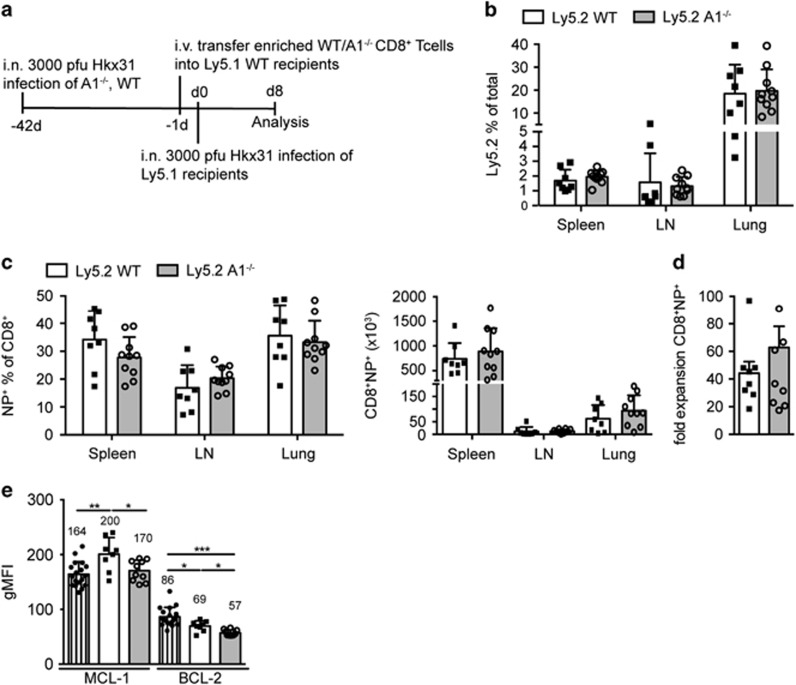
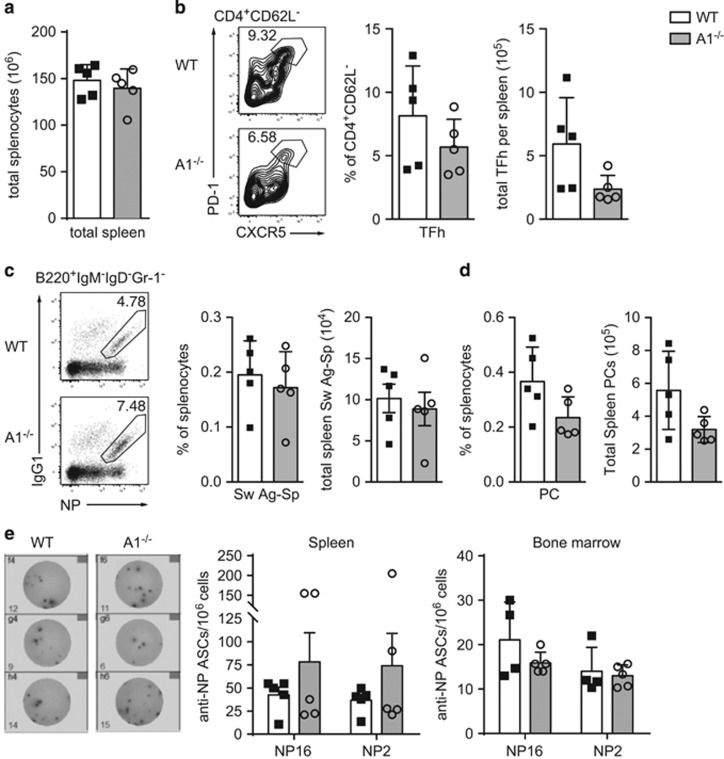
The physiological role of the pro-survival BCL-2 family member A1 has been debated for a long time. Strong mRNA induction in T cells on T cell receptor (TCR)-engagement suggested a major role of A1 in the survival of activated T cells. However, the investigation of the physiological roles of A1 was complicated by the quadruplication of the A1 gene locus in mice, making A1 gene targeting very difficult. Here, we used the recently generated A1-/- mouse model to examine the role of A1 in T cell immunity. We confirmed rapid and strong induction of A1 protein in response to TCR/CD3 stimulation in CD4+ as well as CD8+ T cells. Surprisingly, on infection with the acute influenza HKx31 or the lymphocytic choriomeningitis virus docile strains mice lacking A1 did not show any impairment in the expansion, survival, or effector function of cytotoxic T cells. Furthermore, the ability of A1-/- mice to generate antigen-specific memory T cells or to provide adequate CD4-dependent help to B cells was not impaired. These results suggest functional redundancy of A1 with other pro-survival BCL-2 family members in the control of T cell-dependent immune responses.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Opferman JT, Letai A, Beard C, Sorcinelli MD, Ong CC, Korsmeyer SJ. Development and maintenance of B and T lymphocytes requires antiapoptotic MCL-1. Nature 2003; 426: 671–676. - PubMed
-
- Wensveen FM, van Gisbergen KP, Derks IA, Gerlach C, Schumacher TN, van Lier RA et al. Apoptosis threshold set by Noxa and Mcl-1 after T cell activation regulates competitive selection of high-affinity clones. Immunity 2010; 32: 754–765. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
