

Acylation of the Type 3 Secretion System Translocon Using a Dedicated Acyl Carrier Protein
- PMID: 28085879
- PMCID: PMC5279801
- DOI: 10.1371/journal.pgen.1006556
Acylation of the Type 3 Secretion System Translocon Using a Dedicated Acyl Carrier Protein
Abstract
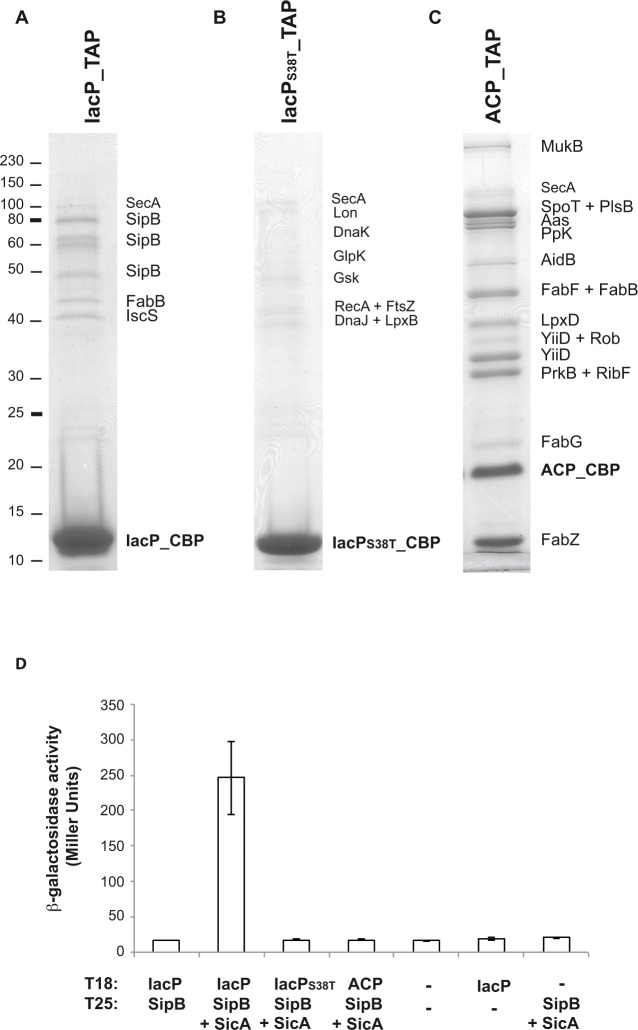
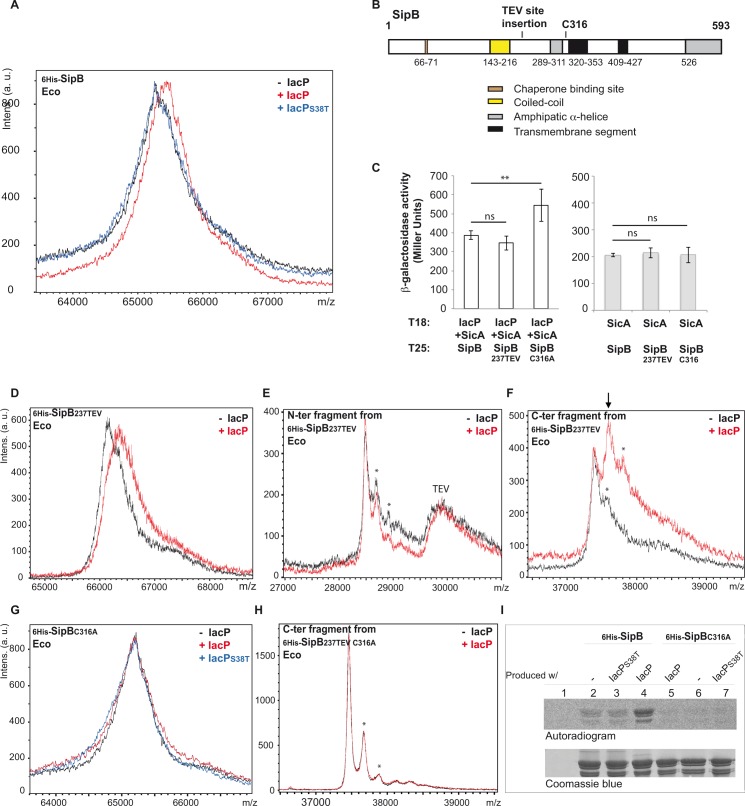
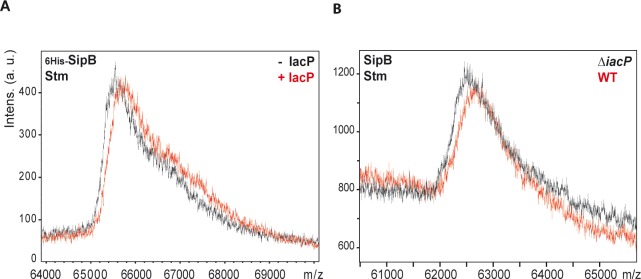
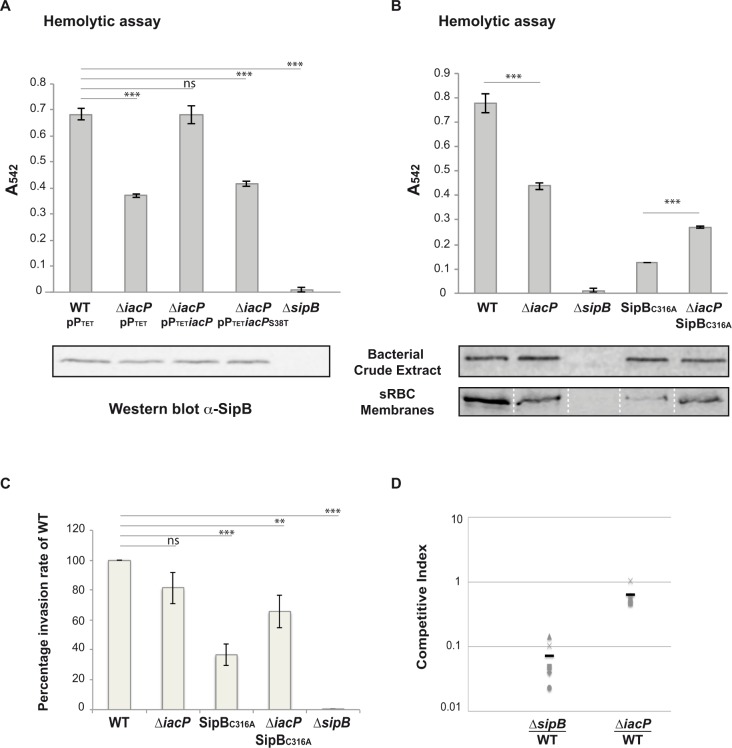
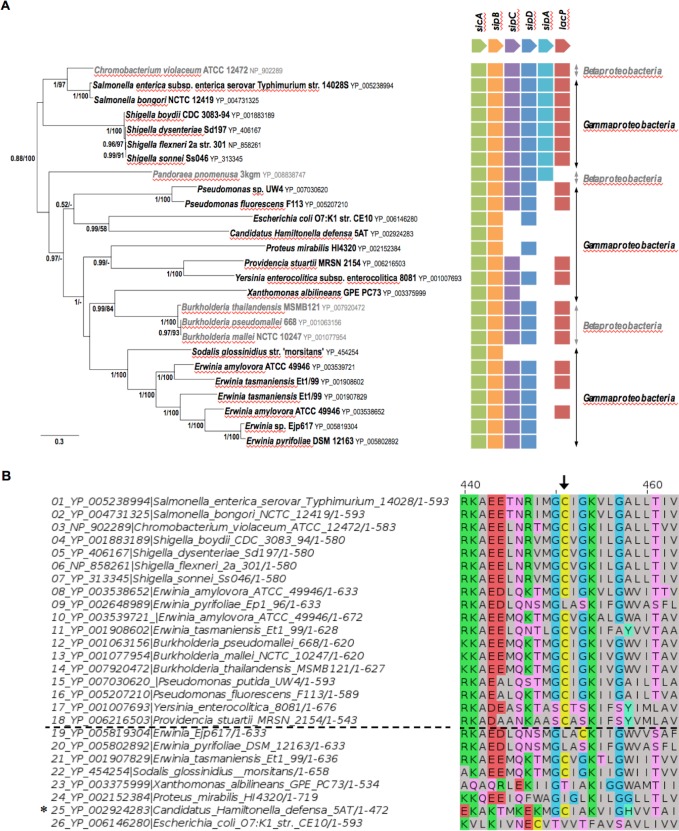
Bacterial pathogens often deliver effectors into host cells using type 3 secretion systems (T3SS), the extremity of which forms a translocon that perforates the host plasma membrane. The T3SS encoded by Salmonella pathogenicity island 1 (SPI-1) is genetically associated with an acyl carrier protein, IacP, whose role has remained enigmatic. In this study, using tandem affinity purification, we identify a direct protein-protein interaction between IacP and the translocon protein SipB. We show, by mass spectrometry and radiolabelling, that SipB is acylated, which provides evidence for a modification of the translocon that has not been described before. A unique and conserved cysteine residue of SipB is identified as crucial for this modification. Although acylation of SipB was not essential to virulence, we show that this posttranslational modification promoted SipB insertion into host-cell membranes and pore-forming activity linked to the SPI-1 T3SS. Cooccurrence of acyl carrier and translocon proteins in several γ- and β-proteobacteria suggests that acylation of the translocon is conserved in these other pathogenic bacteria. These results also indicate that acyl carrier proteins, known for their involvement in metabolic pathways, have also evolved as cofactors of new bacterial protein lipidation pathways.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
