

No Evidence for Phylostratigraphic Bias Impacting Inferences on Patterns of Gene Emergence and Evolution
- PMID: 28087778
- PMCID: PMC5400388
- DOI: 10.1093/molbev/msw284
No Evidence for Phylostratigraphic Bias Impacting Inferences on Patterns of Gene Emergence and Evolution
Abstract
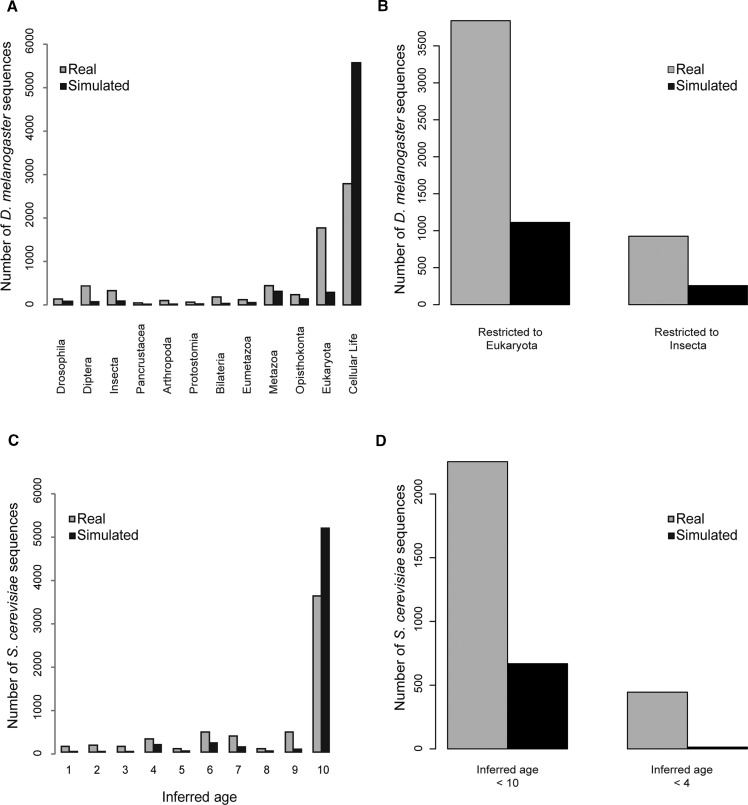
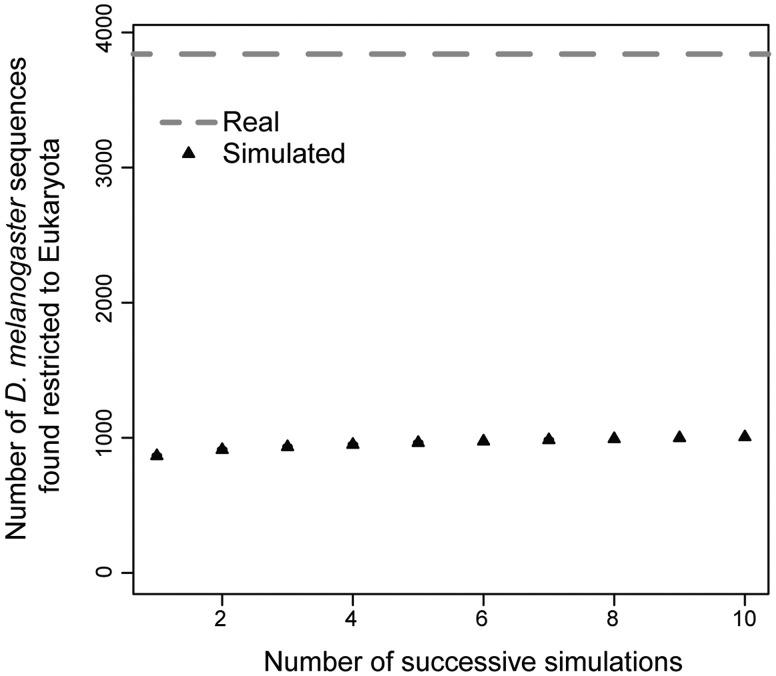
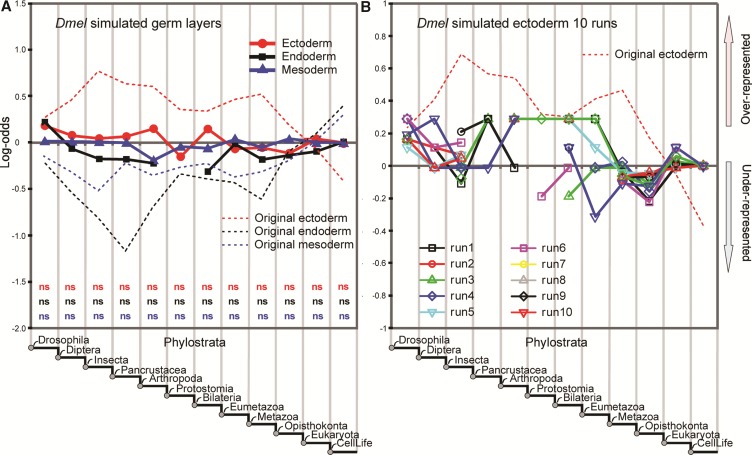
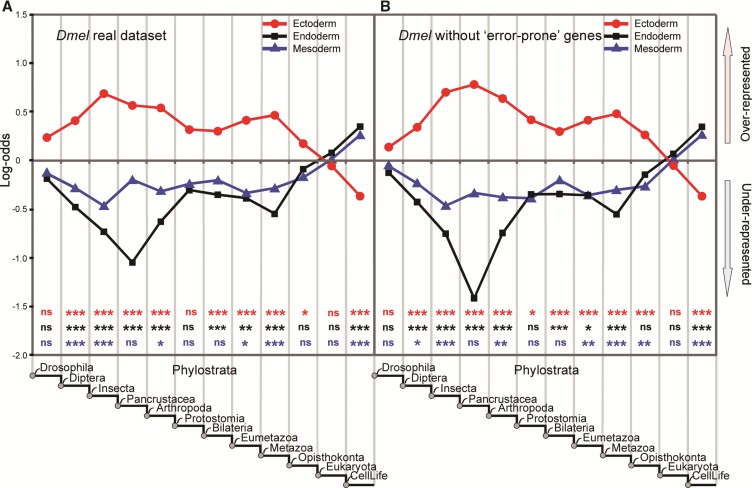
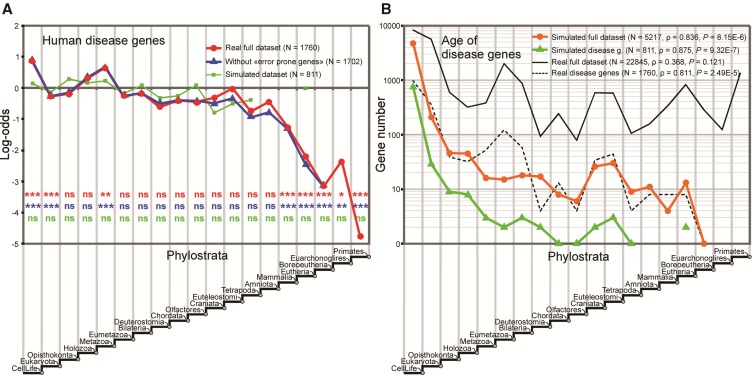
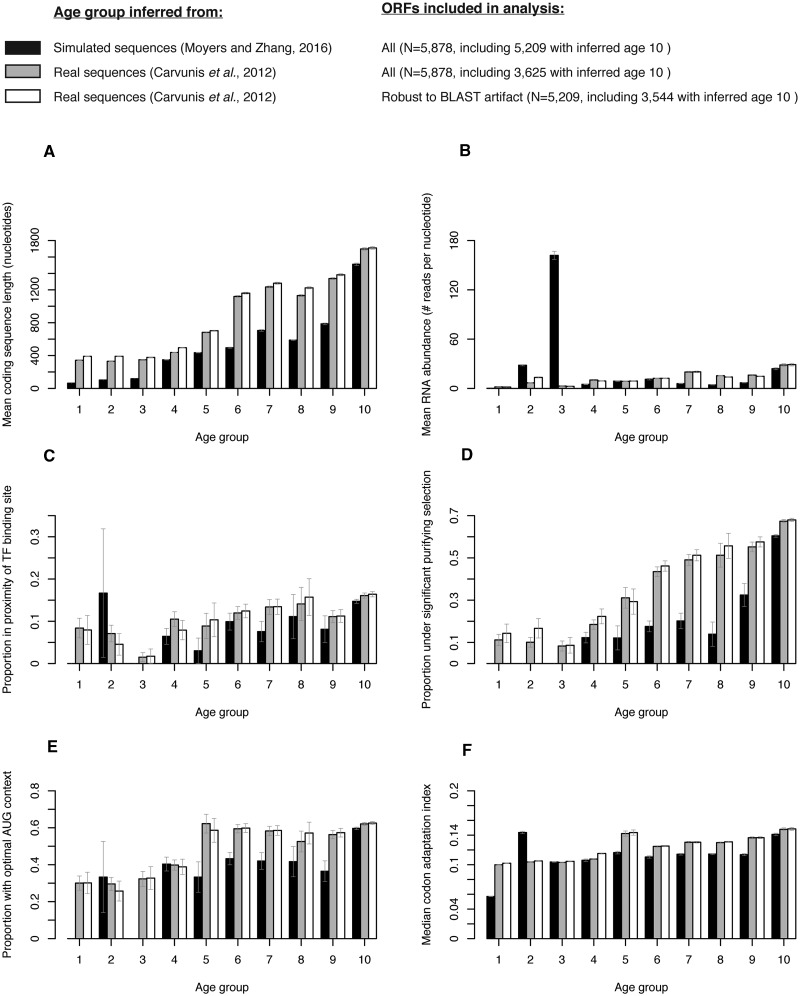
Phylostratigraphy is a computational framework for dating the emergence of DNA and protein sequences in a phylogeny. It has been extensively applied to make inferences on patterns of genome evolution, including patterns of disease gene evolution, ontogeny and de novo gene origination. Phylostratigraphy typically relies on BLAST searches along a species tree, but new simulation studies have raised concerns about the ability of BLAST to detect remote homologues and its impact on phylostratigraphic inferences. Here, we re-assessed these simulations. We found that, even with a possible overall BLAST false negative rate between 11-15%, the large majority of sequences assigned to a recent evolutionary origin by phylostratigraphy is unaffected by technical concerns about BLAST. Where the results of the simulations did cast doubt on previously reported findings, we repeated the original analyses but now excluded all questionable sequences. The originally described patterns remained essentially unchanged. These new analyses strongly support phylostratigraphic inferences, including: genes that emerged after the origin of eukaryotes are more likely to be expressed in the ectoderm than in the endoderm or mesoderm in Drosophila, and the de novo emergence of protein-coding genes from non-genic sequences occurs through proto-gene intermediates in yeast. We conclude that BLAST is an appropriate and sufficiently sensitive tool in phylostratigraphic analysis that does not appear to introduce significant biases into evolutionary pattern inferences.
Keywords: BLAST; gene age estimation; genome analysis; phylostratigraphy.
© The Author 2017. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures

References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
