

Environmental Geometry Aligns the Hippocampal Map during Spatial Reorientation
- PMID: 28089516
- PMCID: PMC5296211
- DOI: 10.1016/j.cub.2016.11.046
Environmental Geometry Aligns the Hippocampal Map during Spatial Reorientation
Abstract
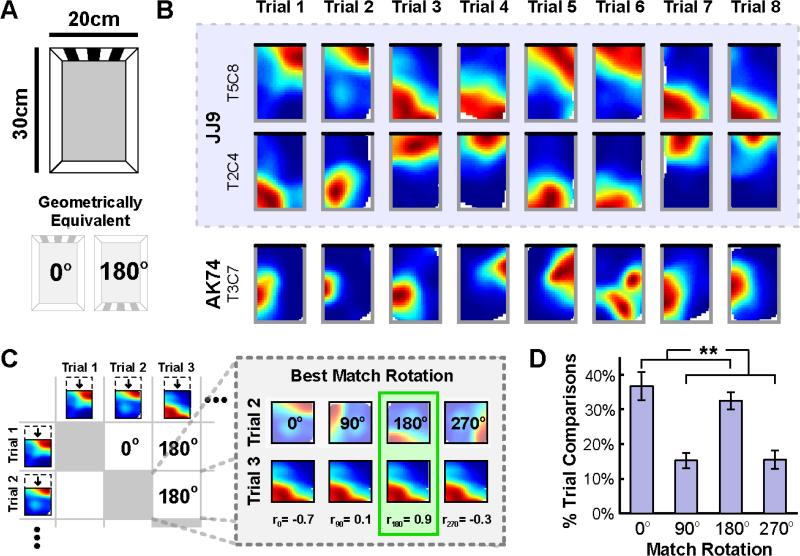
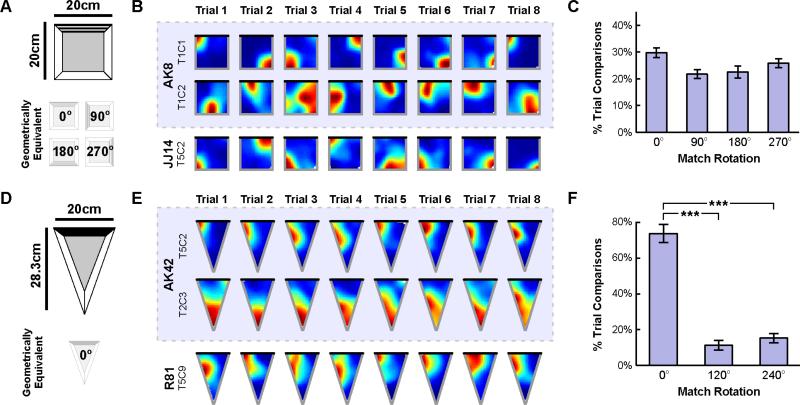
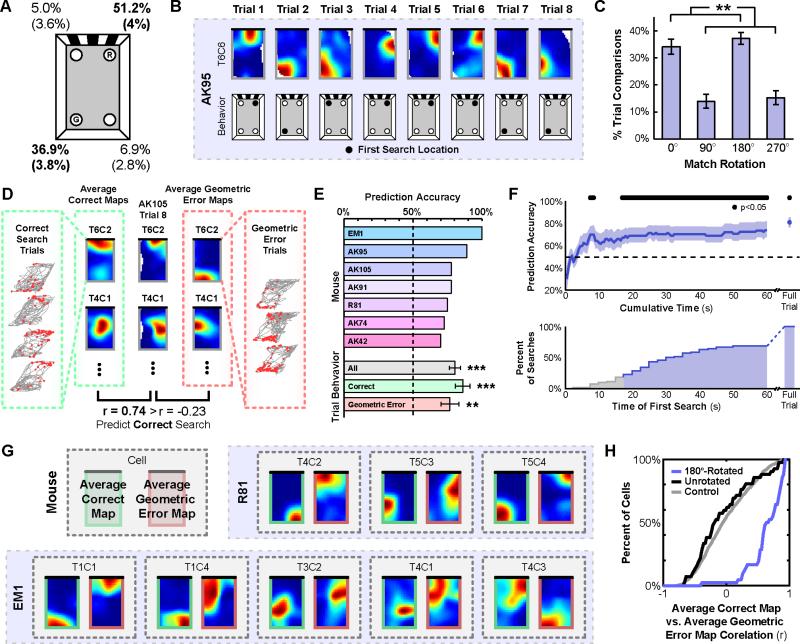
When a navigator's internal sense of direction is disrupted, she must rely on external cues to regain her bearings, a process termed spatial reorientation. Extensive research has demonstrated that the geometric shape of the environment exerts powerful control over reorientation behavior, but the neural and cognitive mechanisms underlying this phenomenon are not well understood. Whereas some theories claim that geometry controls behavior through an allocentric mechanism potentially tied to the hippocampus, others postulate that disoriented navigators reach their goals by using an egocentric view-matching strategy. To resolve this debate, we characterized hippocampal representations during reorientation. We first recorded from CA1 cells as disoriented mice foraged in chambers of various shapes. We found that the alignment of the recovered hippocampal map was determined by the geometry of the chamber, but not by nongeometric cues, even when these cues could be used to disambiguate geometric ambiguities. We then recorded hippocampal activity as disoriented mice performed a classical goal-directed spatial memory task in a rectangular chamber. Again, we found that the recovered hippocampal map aligned solely to the chamber geometry. Critically, we also found a strong correspondence between the hippocampal map alignment and the animal's behavior, making it possible to predict the search location of the animal from neural responses on a trial-by-trial basis. Together, these results demonstrate that spatial reorientation involves the alignment of the hippocampal map to local geometry. We hypothesize that geometry may be an especially salient cue for reorientation because it is an inherently stable aspect of the environment.
Keywords: cognitive map; disorientation; geometric module; hippocampus; navigation; place cells; spatial geometry; spatial reorientation.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
Comment in
-
Navigation: Whence Our Sense of Direction?Curr Biol. 2017 Feb 6;27(3):R108-R110. doi: 10.1016/j.cub.2016.11.044. Curr Biol. 2017. PMID: 28171754
References
-
- Mittelstaedt M-L, Mittelstaedt H. Homing by path integration in a mammal. Naturwissenschaften. 1980;67:566–567.
-
- Cheng K, Huttenlocher J, Newcombe NS. 25 years of research on the use of geometry in spatial reorientation: a current theoretical perspective. Psychon. Bull. Rev. 2013;20:1033–54. - PubMed
-
- Cheng K, Newcombe NS. Is there a geometric module for spatial orientation? Squaring theory and evidence. Psychon. Bull. Rev. 2005;12:1–23. - PubMed
-
- Cheng K. A purely geometric module in the rat's spatial representation. Cognition. 1986;23:149–78. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
