

Prevention of Dietary-Fat-Fueled Ketogenesis Attenuates BRAF V600E Tumor Growth
- PMID: 28089569
- PMCID: PMC5299059
- DOI: 10.1016/j.cmet.2016.12.010
Prevention of Dietary-Fat-Fueled Ketogenesis Attenuates BRAF V600E Tumor Growth
Abstract
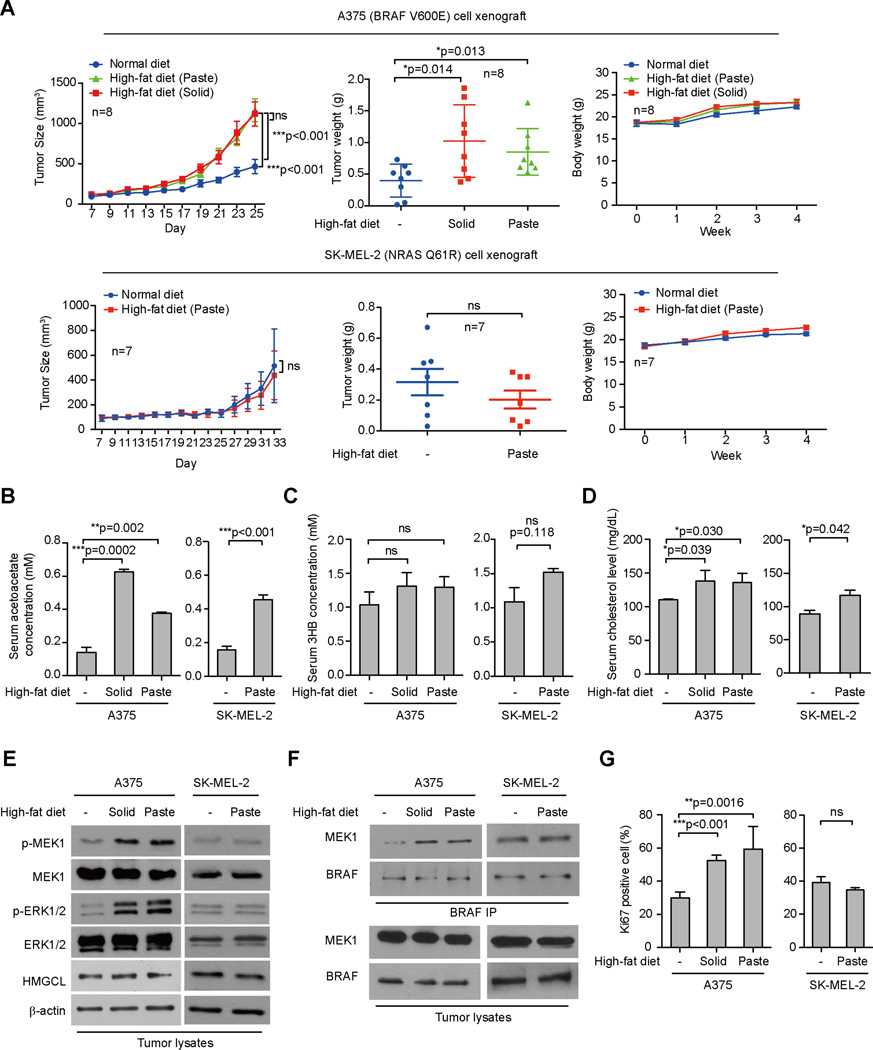
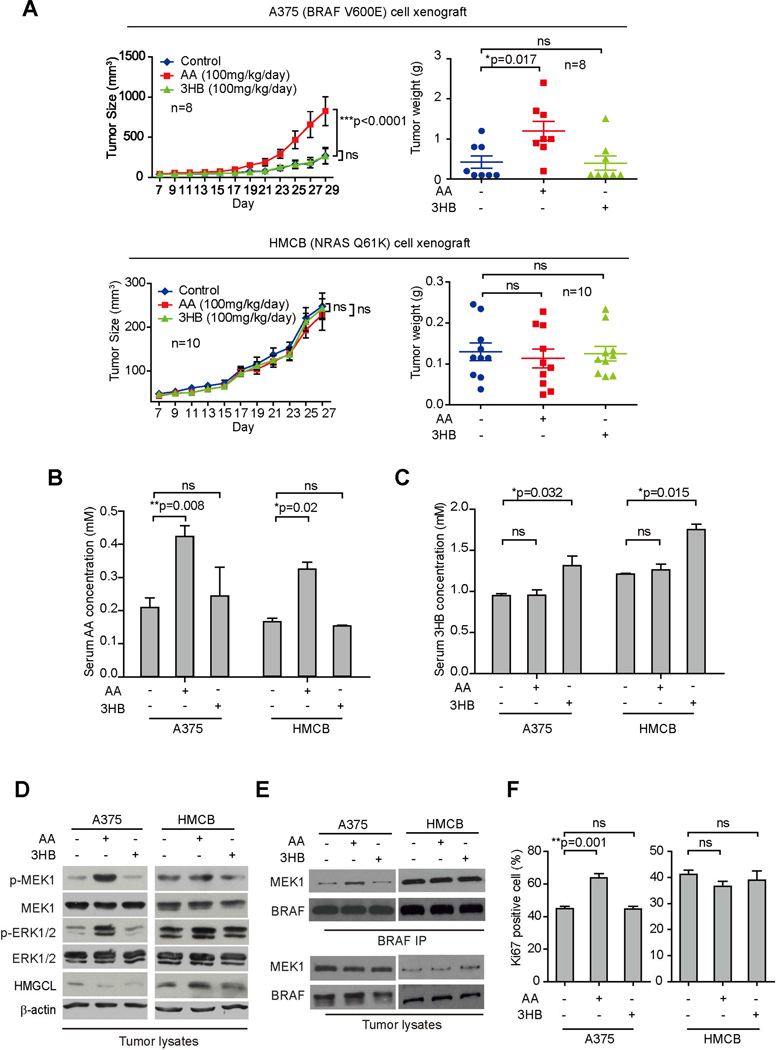
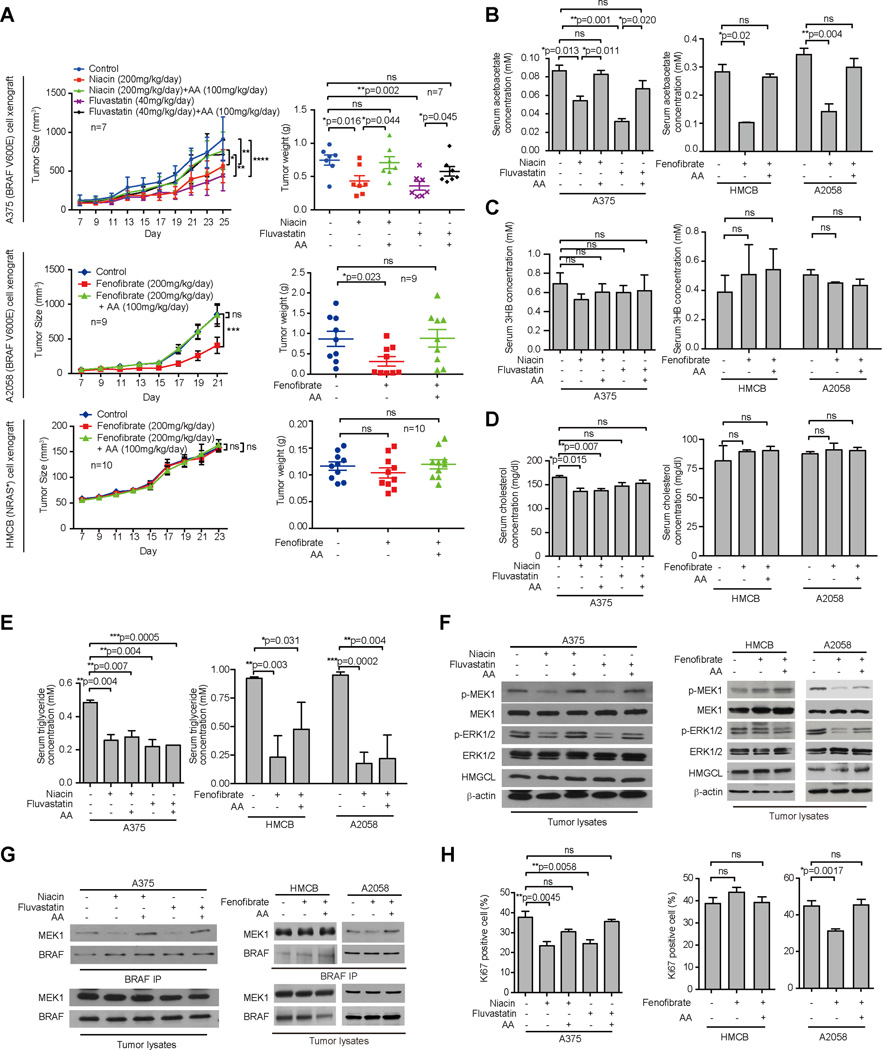
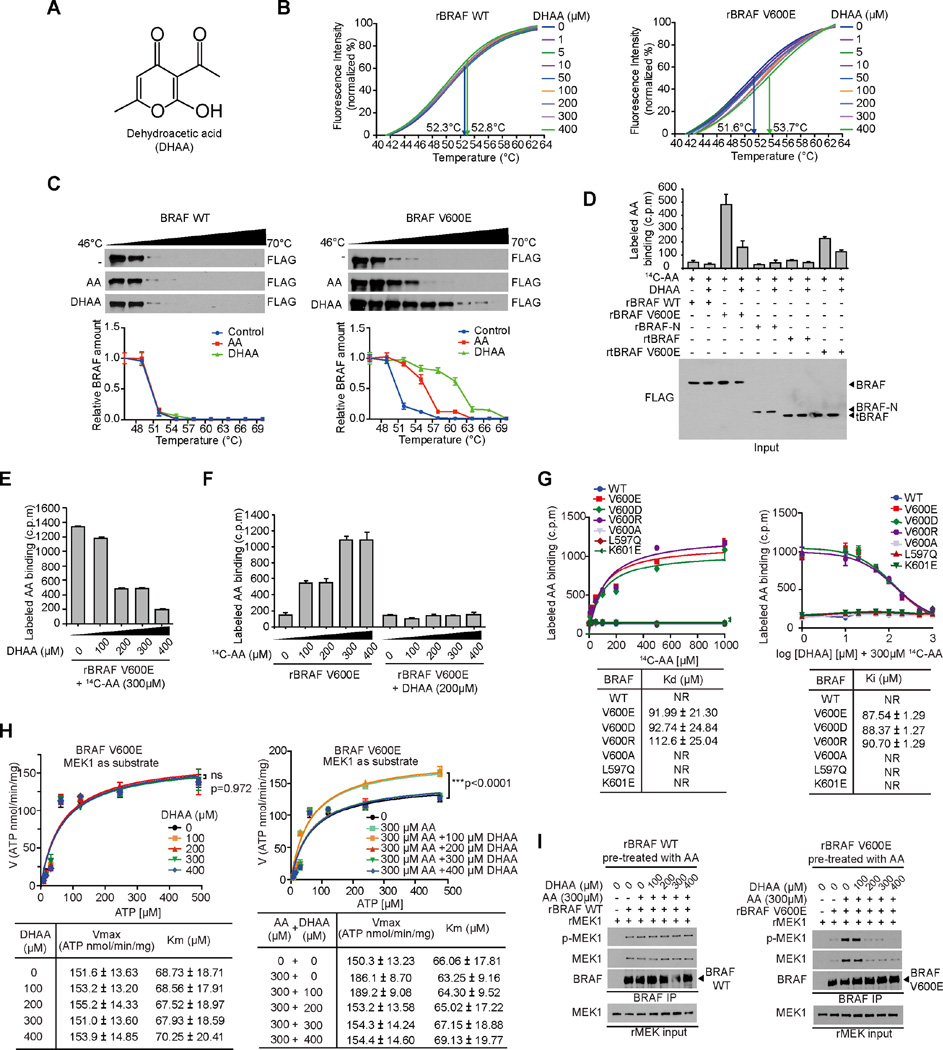
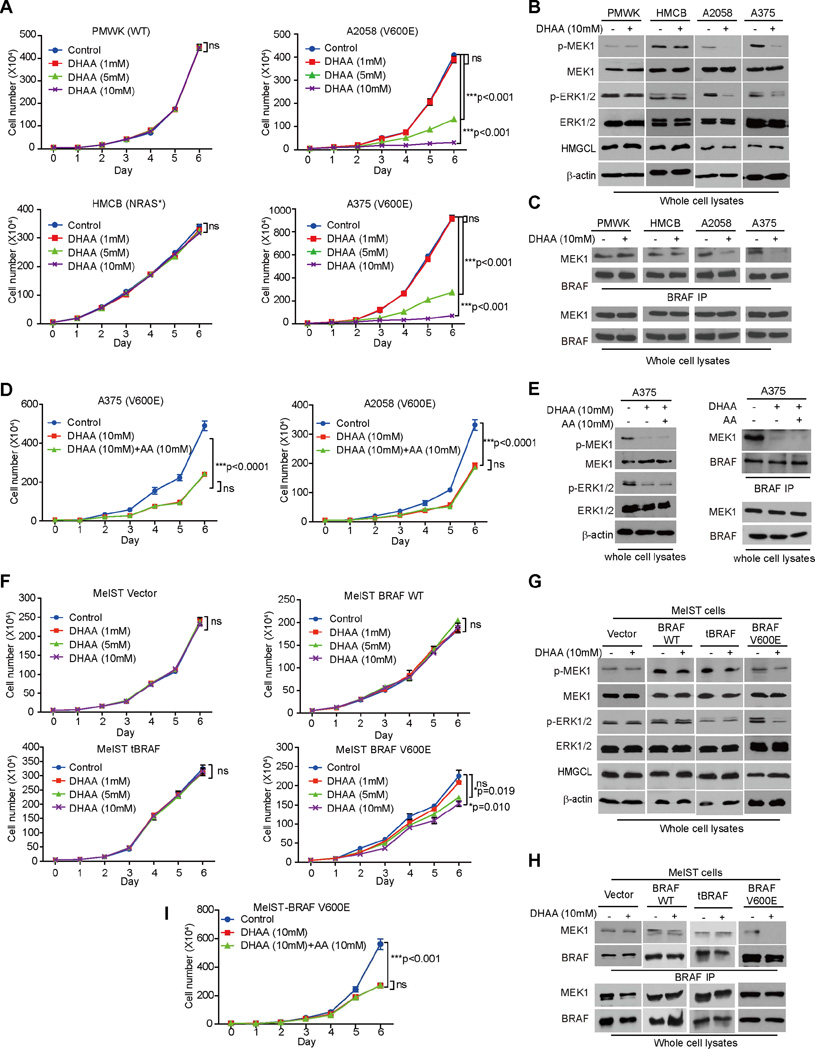
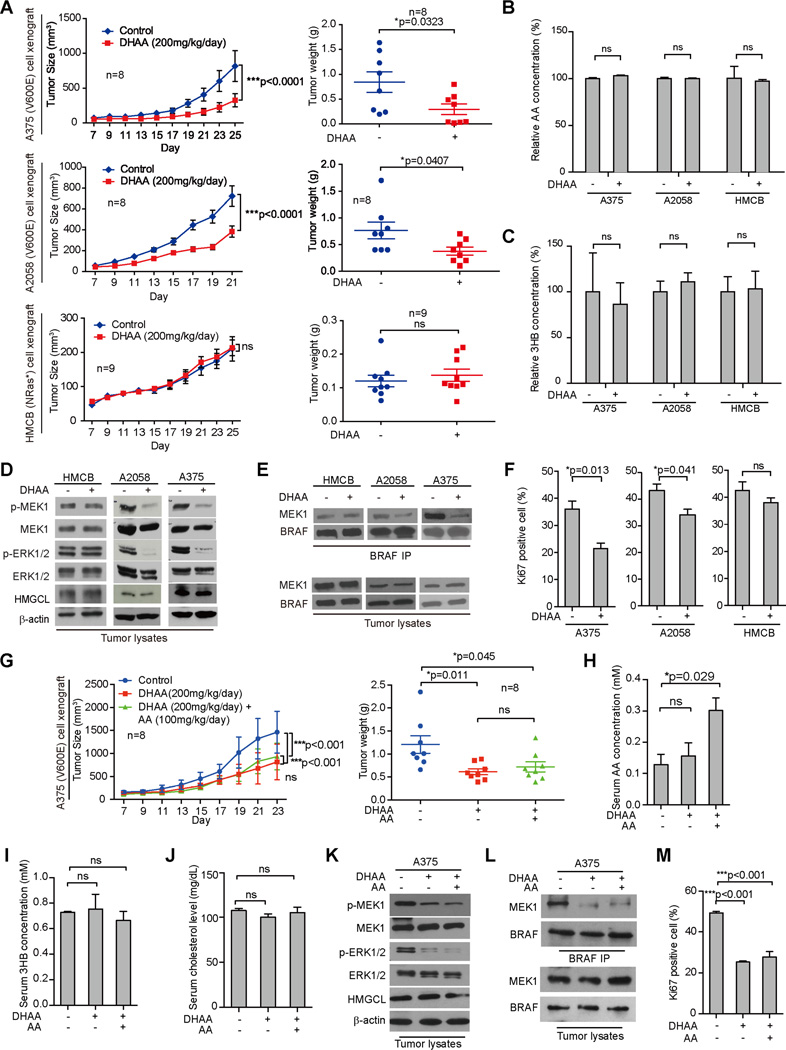
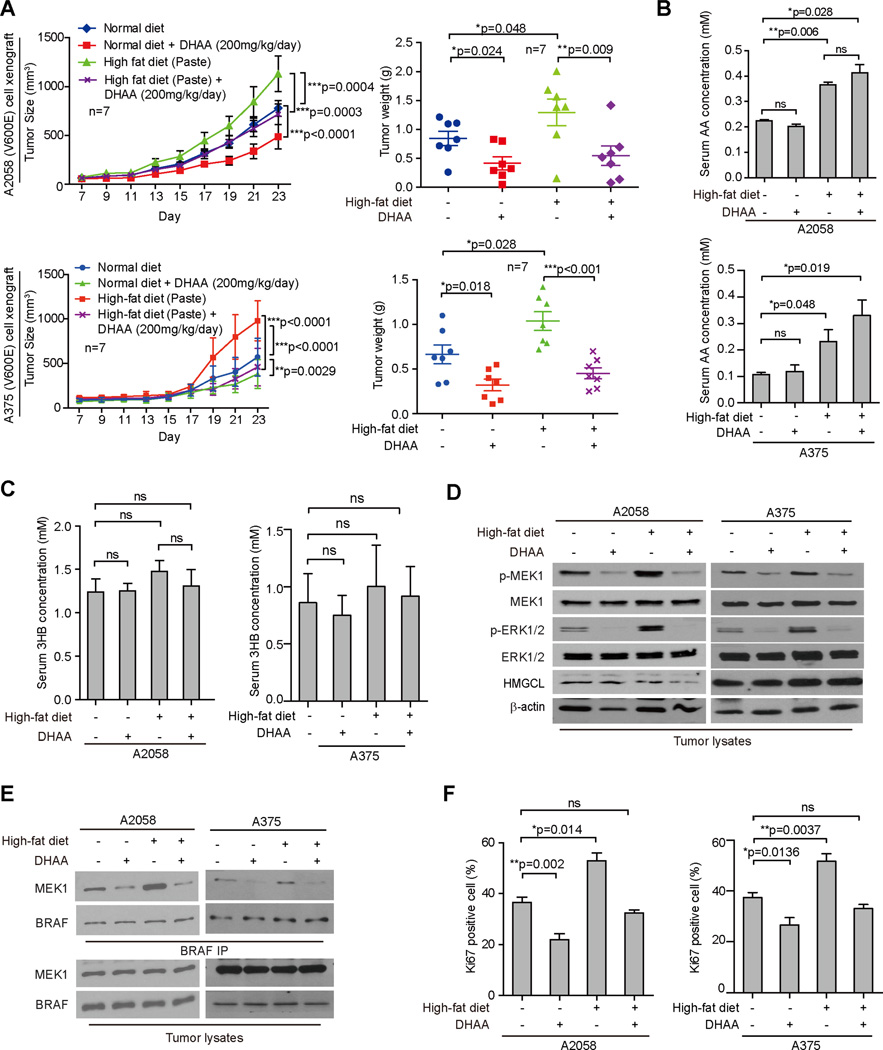
Lifestyle factors, including diet, play an important role in the survival of cancer patients. However, the molecular mechanisms underlying pathogenic links between diet and particular oncogenic mutations in human cancers remain unclear. We recently reported that the ketone body acetoacetate selectively enhances BRAF V600E mutant-dependent MEK1 activation in human cancers. Here we show that a high-fat ketogenic diet increased serum levels of acetoacetate, leading to enhanced tumor growth potential of BRAF V600E-expressing human melanoma cells in xenograft mice. Treatment with hypolipidemic agents to lower circulating acetoacetate levels or an inhibitory homolog of acetoacetate, dehydroacetic acid, to antagonize acetoacetate-BRAF V600E binding attenuated BRAF V600E tumor growth. These findings reveal a signaling basis underlying a pathogenic role of dietary fat in BRAF V600E-expressing melanoma, providing insights into the design of conceptualized "precision diets" that may prevent or delay tumor progression based on an individual's specific oncogenic mutation profile.
Keywords: BRAF V600E; acetoacetate; cancer metabolism; cancer prevention; cancer risk; cancer therapy; dehydroacetic acid; dietary fat; ketogenesis; precision diet.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Antoniadis AG, Petridou ET, Antonopoulos CN, Dessypris N, Panagopoulou P, Chamberland JP, Adami HO, Gogas H, Mantzoros CS. Insulin resistance in relation to melanoma risk. Melanoma Res. 2011;21:541–546. - PubMed
-
- Arcaini L, Zibellini S, Boveri E, Riboni R, Rattotti S, Varettoni M, Guerrera ML, Lucioni M, Tenore A, Merli M, Rizzi S, Morello L, Cavalloni C, Da Via MC, Paulli M, Cazzola M. The BRAF V600E mutation in hairy cell leukemia and other mature B-cell neoplasms. Blood. 2012;119:188–191. - PubMed
-
- Avis HJ, Vissers MN, Stein EA, Wijburg FA, Trip MD, Kastelein JJ, Hutten BA. A systematic review and meta-analysis of statin therapy in children with familial hypercholesterolemia. Arterioscler Thromb Vasc Biol. 2007;27:1803–1810. - PubMed
-
- Balasse EO, Fery F. Ketone body production and disposal: effects of fasting, diabetes, and exercise. Diabetes Metab Rev. 1989;5:247–270. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
