

A disrupted transsulphuration pathway results in accumulation of redox metabolites and induction of gametocytogenesis in malaria
- PMID: 28091526
- PMCID: PMC5238400
- DOI: 10.1038/srep40213
A disrupted transsulphuration pathway results in accumulation of redox metabolites and induction of gametocytogenesis in malaria
Abstract
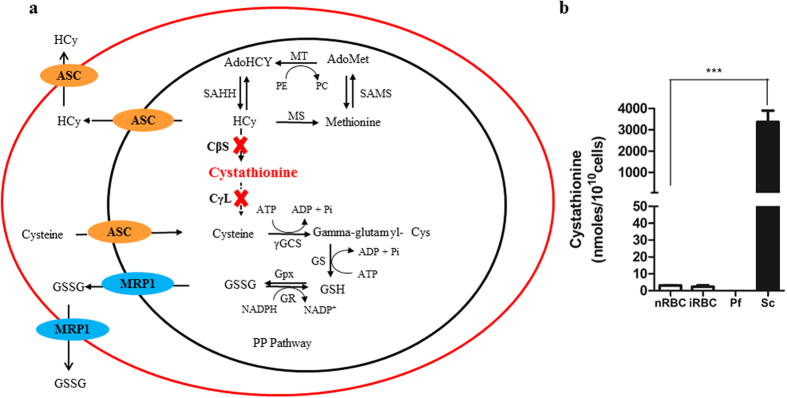
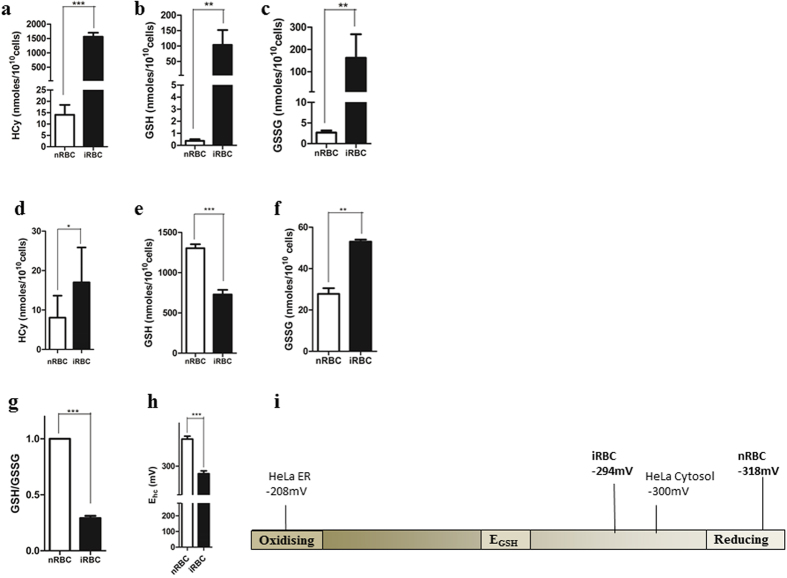
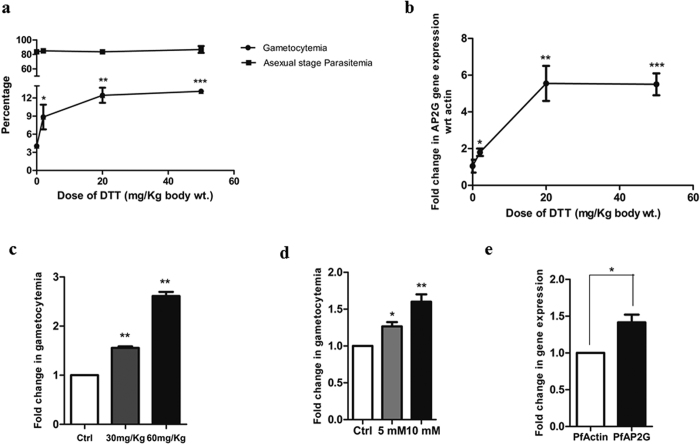
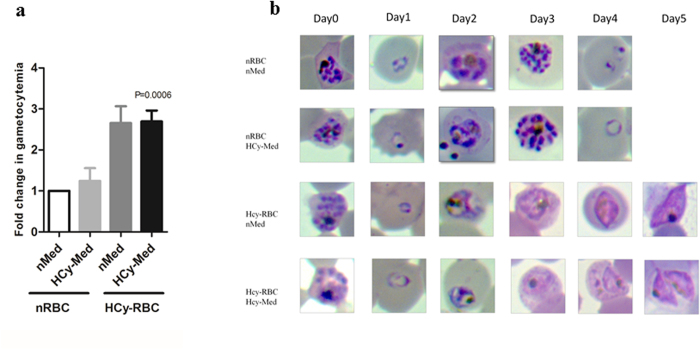
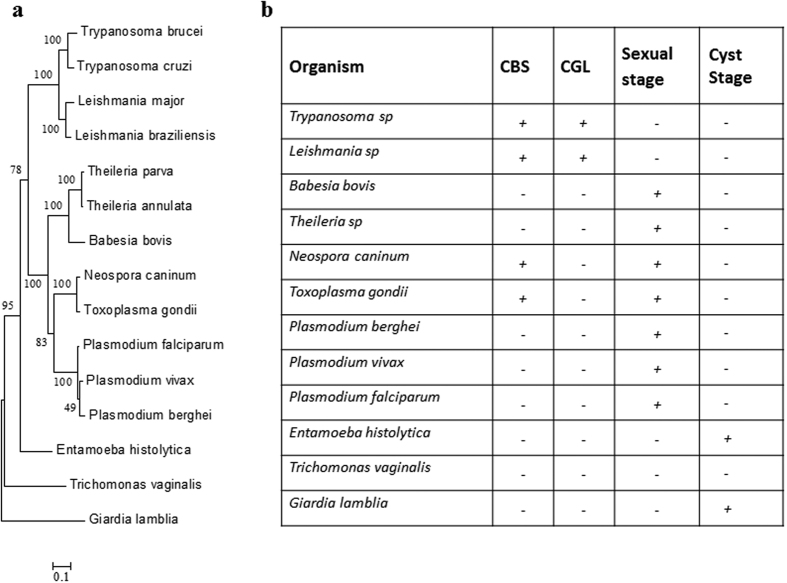
Intra-erythrocytic growth of malaria parasite is known to induce redox stress. In addition to haem degradation which generates reactive oxygen species (ROS), the parasite is also thought to efflux redox active homocysteine. To understand the basis underlying accumulation of homocysteine, we have examined the transsulphuration (TS) pathway in the parasite, which is known to convert homocysteine to cysteine in higher eukaryotes. Our bioinformatic analysis revealed absence of key enzymes in the biosynthesis of cysteine namely cystathionine-β-synthase and cystathionine-γ-lyase in the parasite. Using mass spectrometry, we confirmed the absence of cystathionine, which is formed by enzymatic conversion of homocysteine thereby confirming truncation of TS pathway. We also quantitated levels of glutathione and homocysteine in infected erythrocytes and its spent medium. Our results showed increase in levels of these metabolites intracellularly and in culture supernatants. Our results provide a mechanistic basis for the long-known occurrence of hyperhomocysteinemia in malaria. Most importantly we find that homocysteine induces the transcription factor implicated in gametocytogenesis namely AP2-G and consequently triggers sexual stage conversion. We confirmed this observation both in vitro using Plasmodium falciparum cultures, and in vivo in the mouse model of malaria. Our study implicates homocysteine as a potential physiological trigger of gametocytogenesis.
Figures
References
-
- Atamna H. & Ginsburg H. The malaria parasite supplies glutathione to its host cell–investigation of glutathione transport and metabolism in human erythrocytes infected with Plasmodium falciparum. Eur J Biochem 250, 670–679 (1997). - PubMed
-
- Vennerstrom J. L. & Eaton J. W. Oxidants, oxidant drugs, and malaria. J Med Chem 31, 1269–1277 (1988). - PubMed
-
- Jortzik E. & Becker K. Thioredoxin and glutathione systems in Plasmodium falciparum. Int J Med Microbiol 302, 187–194, doi: S1438-4221(12)00035-5 (2012). - PubMed
-
- Kawazu S., Komaki-Yasuda K., Oku H. & Kano S. Peroxiredoxins in malaria parasites: parasitologic aspects. Parasitol Int 57, 1–7, doi: S1383-5769(07)00101-8 (2008). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
