

Effects of forced movements on learning: Findings from a choice reaction time task in rats
- PMID: 28091814
- PMCID: PMC5429386
- DOI: 10.3758/s13420-016-0255-9
Effects of forced movements on learning: Findings from a choice reaction time task in rats
Abstract
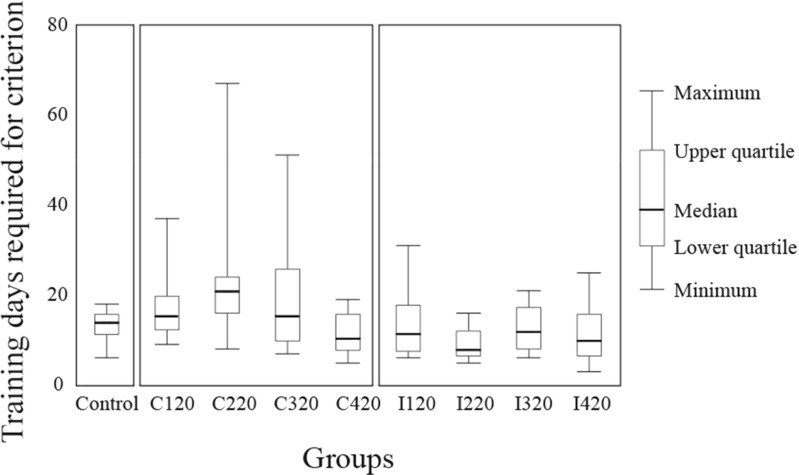
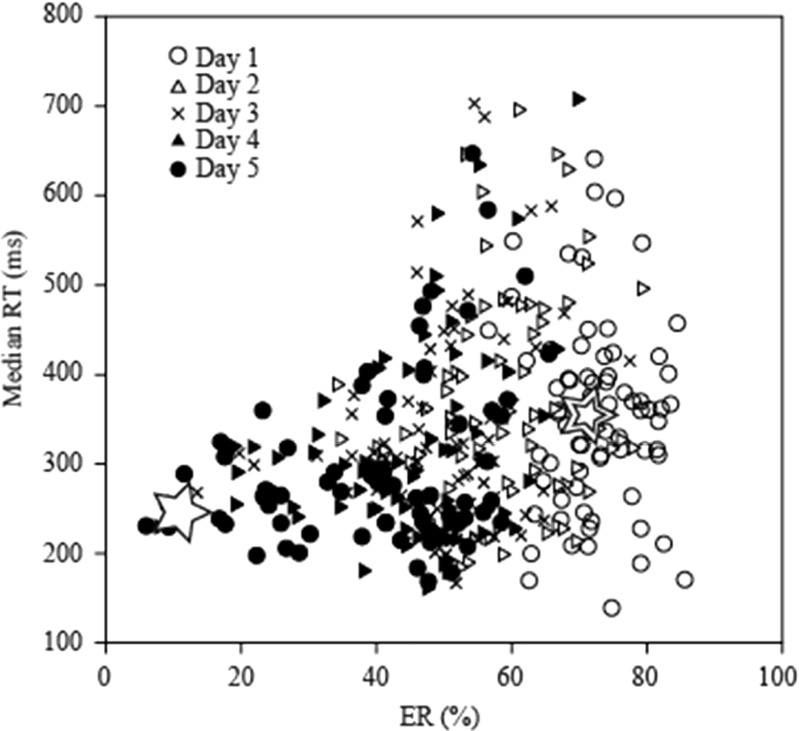
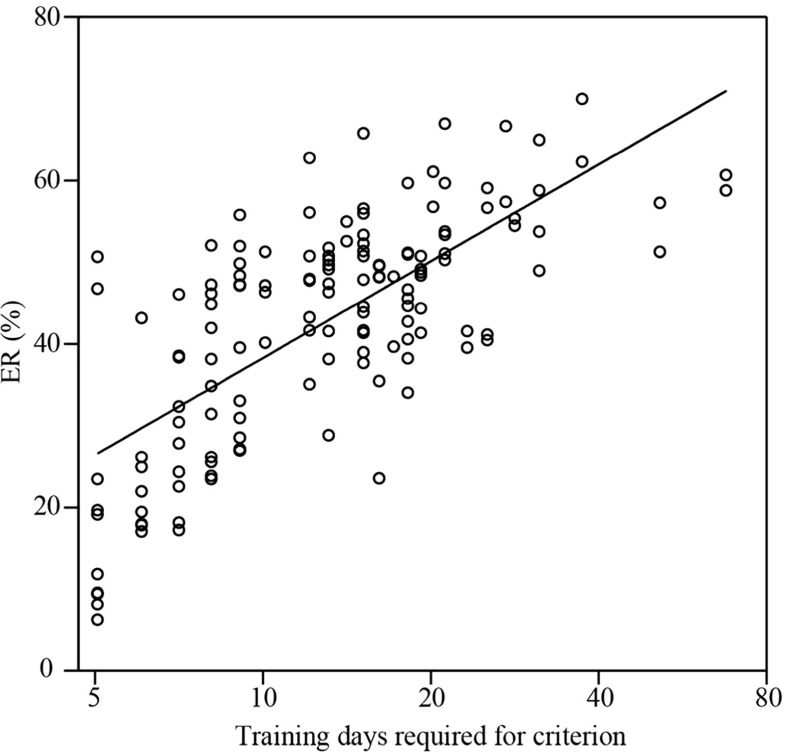
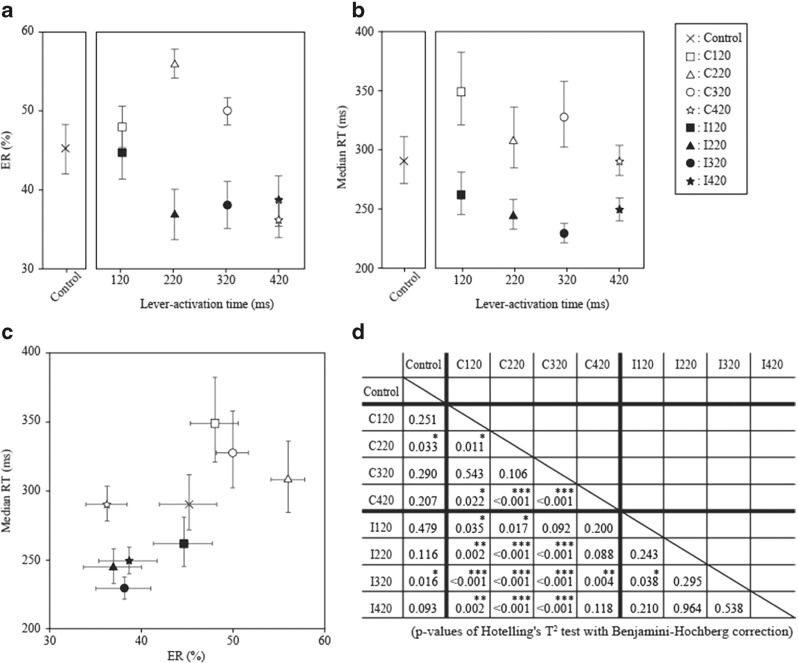
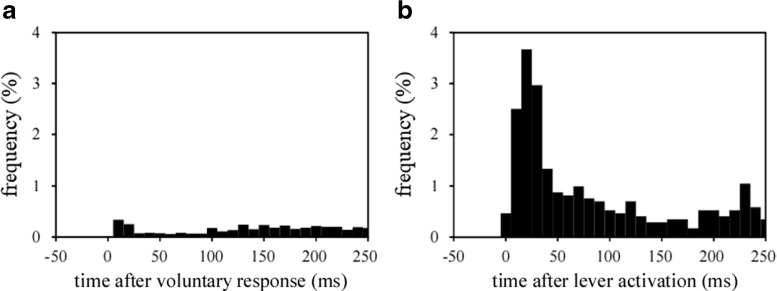
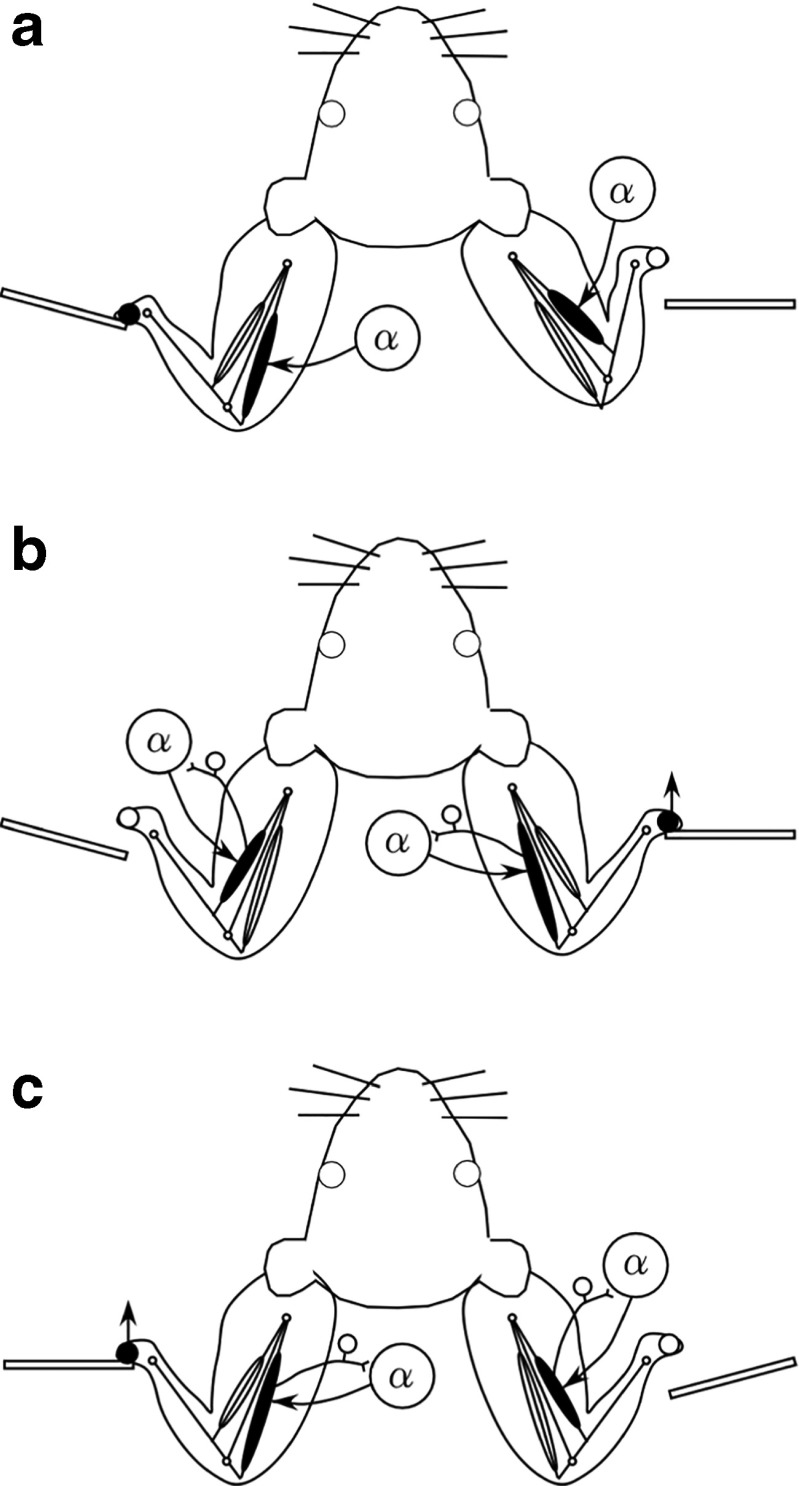
To investigate how motor sensation facilitates learning, we used a sensory-motor association task to determine whether the sensation induced by forced movements contributes to performance improvements in rats. The rats were trained to respond to a tactile stimulus (an air puff) by releasing a lever pressed by the stimulated (compatible condition) or nonstimulated (incompatible condition) forepaw. When error rates fell below 15%, the compatibility condition was changed (reversal learning). An error trial was followed by a lever activation trial in which a lever on the correct or the incorrect response side was automatically elevated at a preset time of 120, 220, 320, or 420 ms after tactile stimulation. This lever activation induced forepaw movement similar to that in a voluntary lever release response, and also induced body movement that occasionally caused elevation of the other forepaw. The effects of lever activation may have produced a sensation similar to that of voluntary lever release by the forepaw on the nonactivated lever. We found that the performance improvement rate was increased by the lever activation procedure on the incorrect response side (i.e., with the nonactivated lever on the correct response side). Furthermore, the performance improvement rate changed depending on the timing of lever activation: Facilitative effects were largest with lever activation on the incorrect response side at 320 ms after tactile stimulation, whereas hindering effects were largest for lever activation on the correct response side at 220 ms after tactile stimulation. These findings suggest that forced movements, which provide tactile and proprioceptive stimulation, affect sensory-motor associative learning in a time-dependent manner.
Keywords: Actuator; Behavior; Proprioceptive sensation; Robotics; Voluntary movement.
Figures
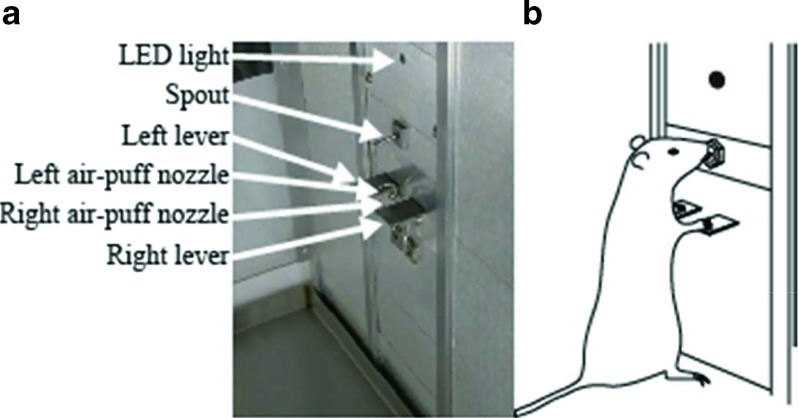
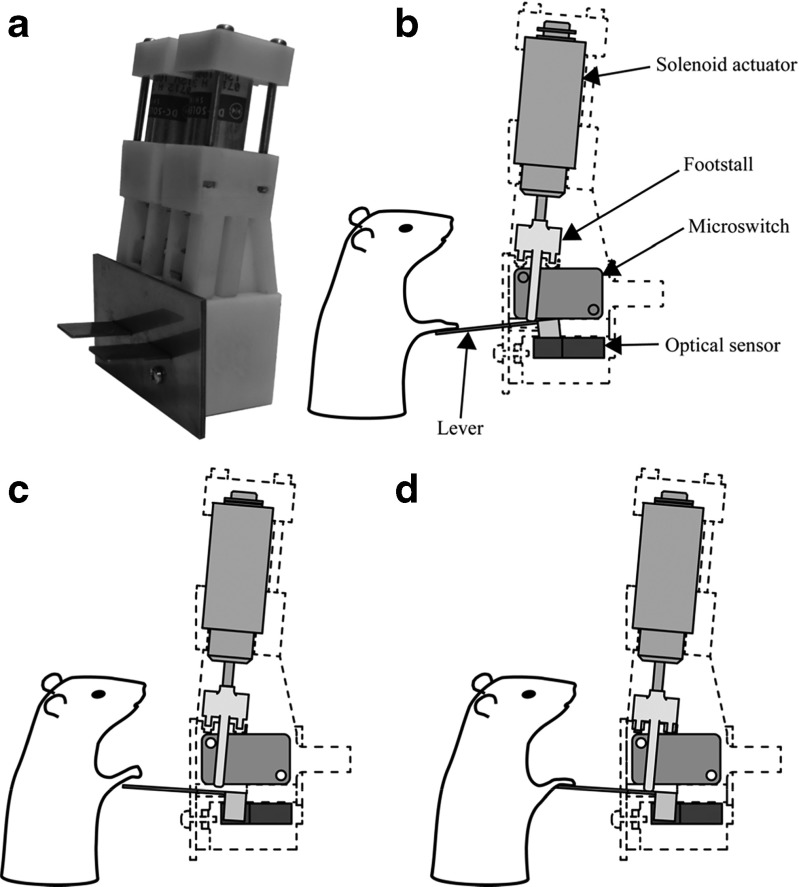
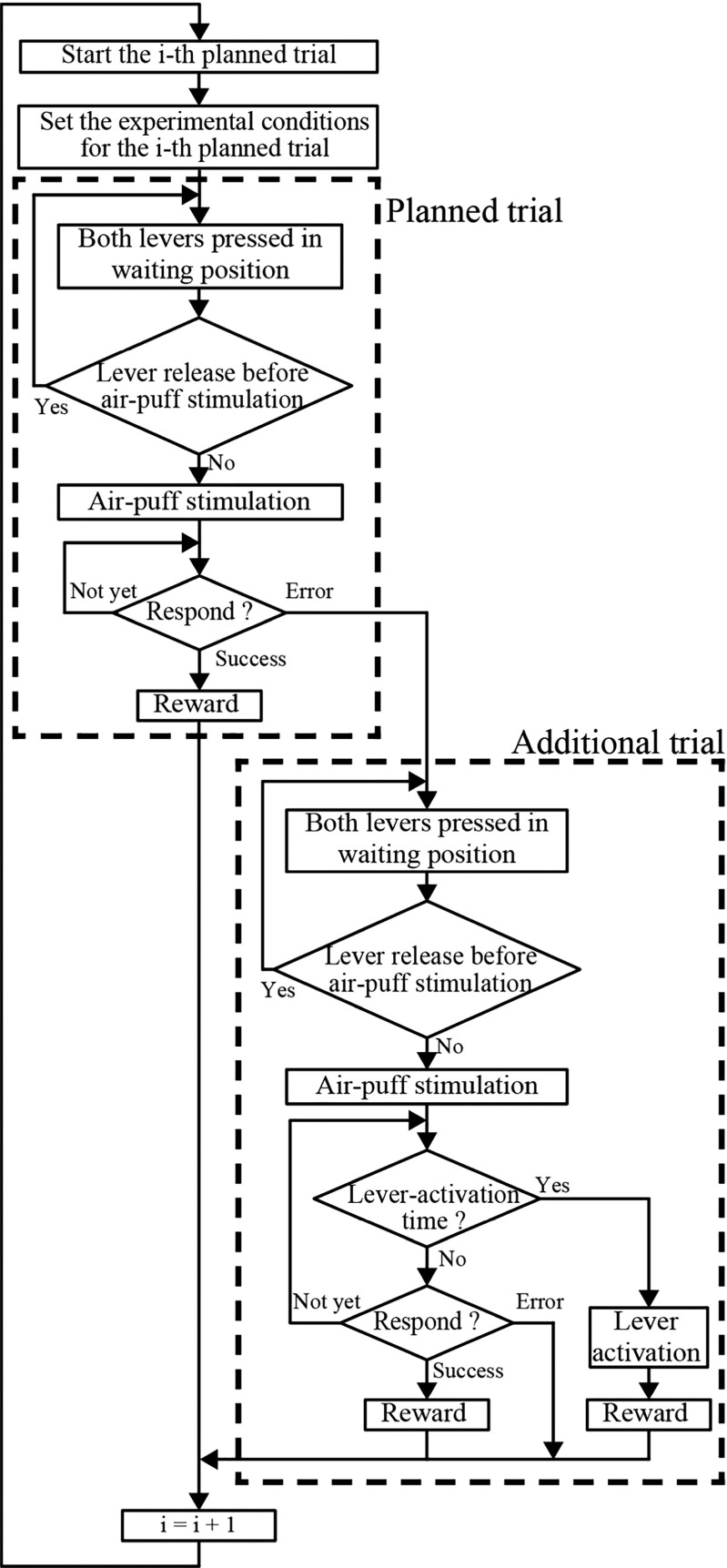
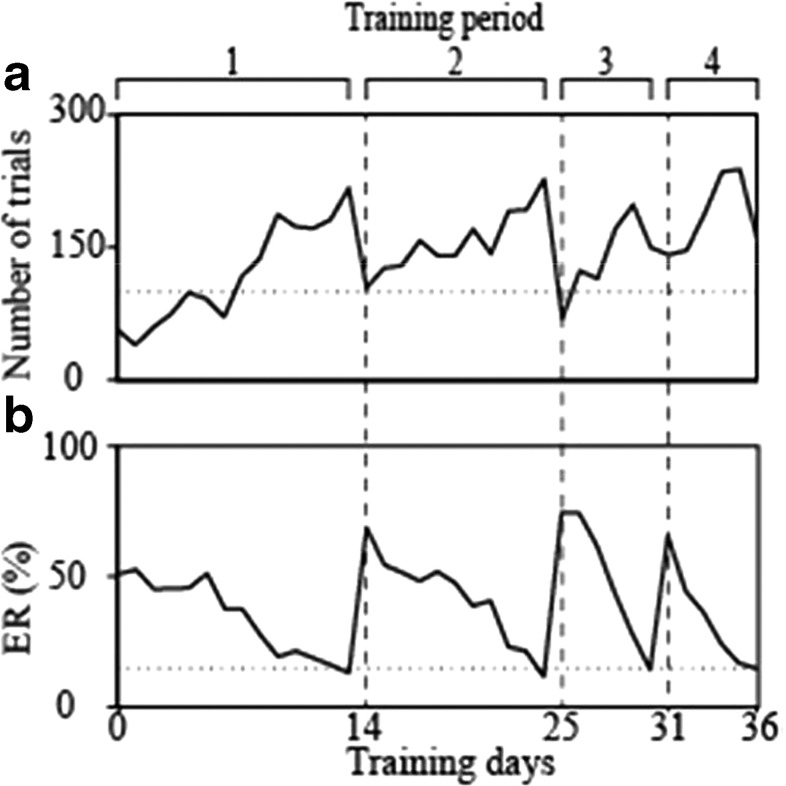
References
-
- Benjamini Y, Hochberg Y. Controlling the false discovery rate—A practical and powerful approach to multiple testing. Journal of the Royal Statistical Society: Series B. 1995;57:289–300.
-
- Boudreau MJ, Smith AM. Activity in rostral motor cortex in response to predictable force-pulse perturbations in a precision grip task. Journal of Neurophysiology. 2001;86:1079–1085. - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
