

Vocal development in a Waddington landscape
- PMID: 28092262
- PMCID: PMC5310845
- DOI: 10.7554/eLife.20782
Vocal development in a Waddington landscape
Abstract
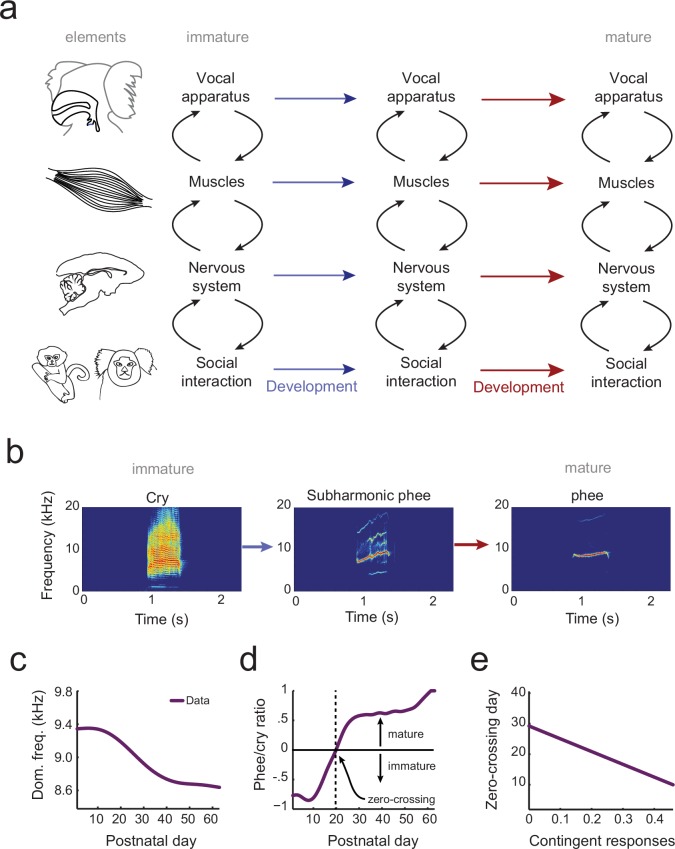
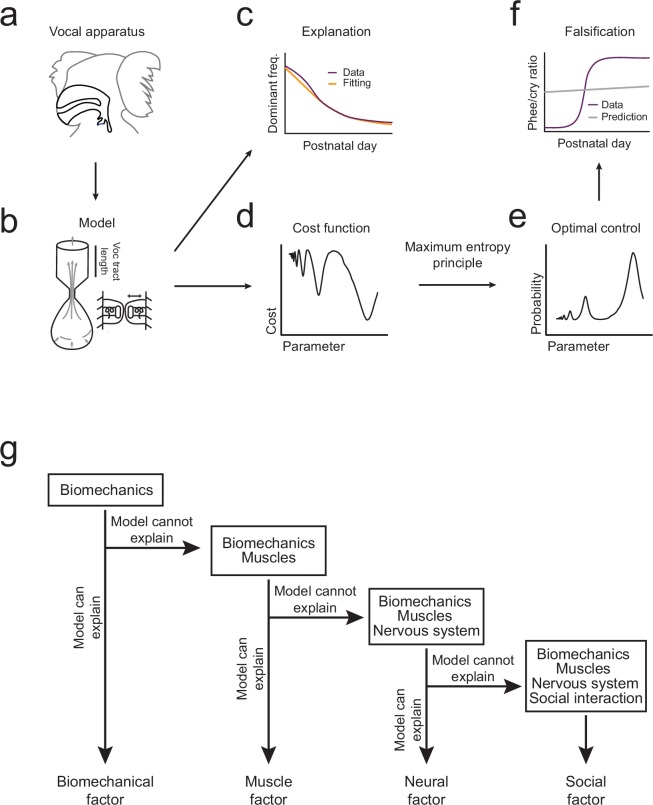
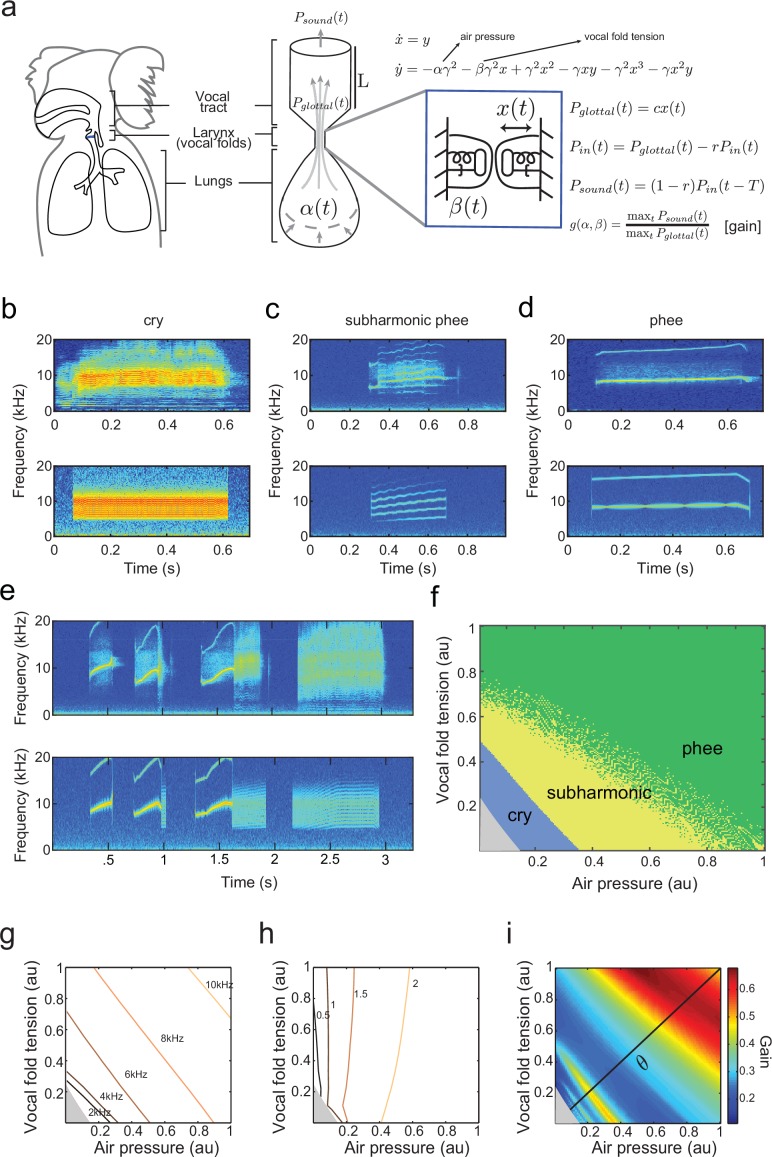
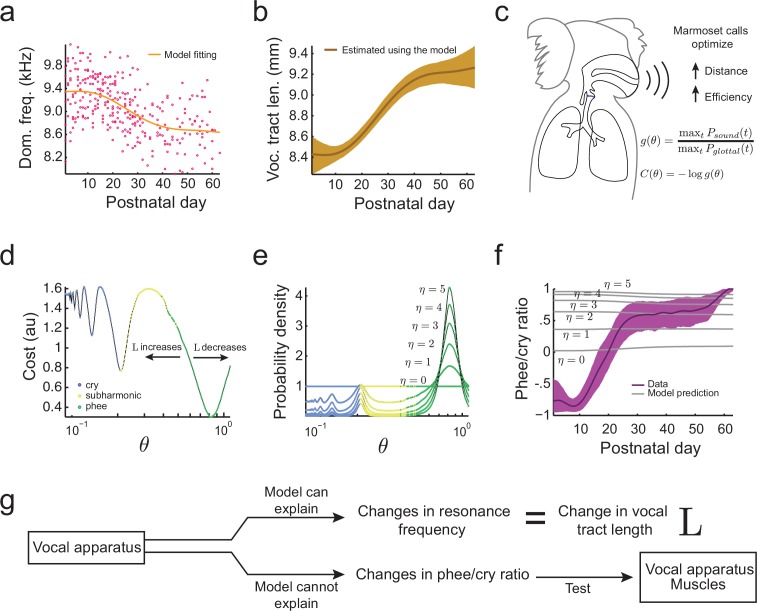
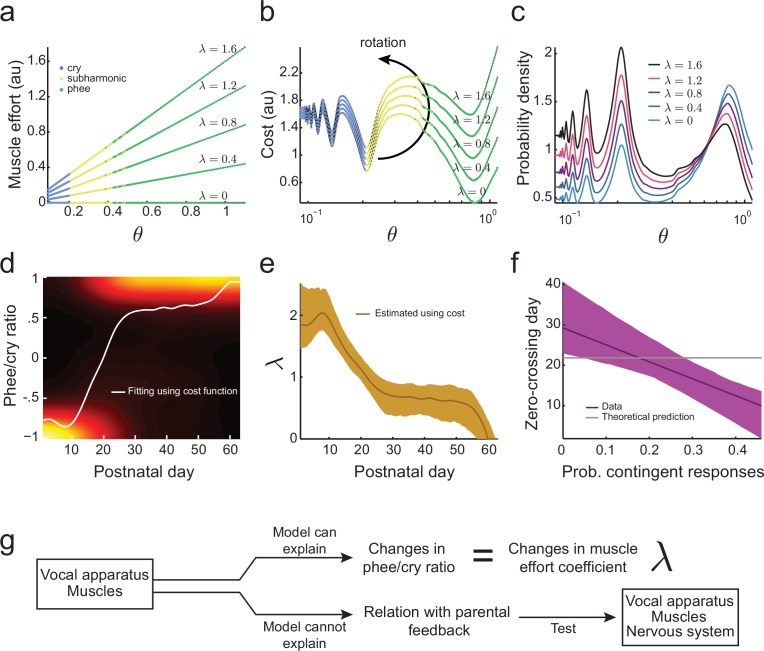
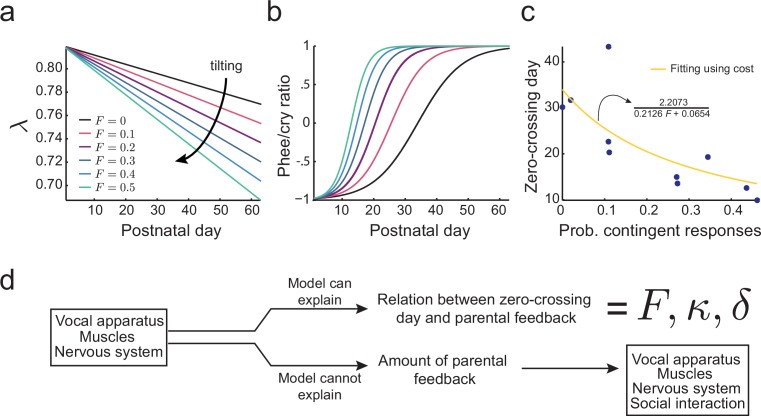
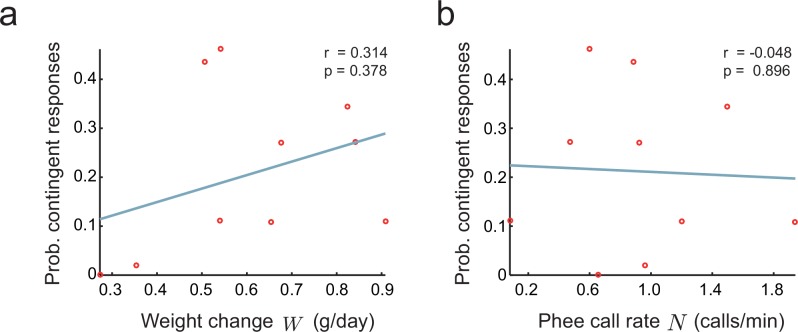
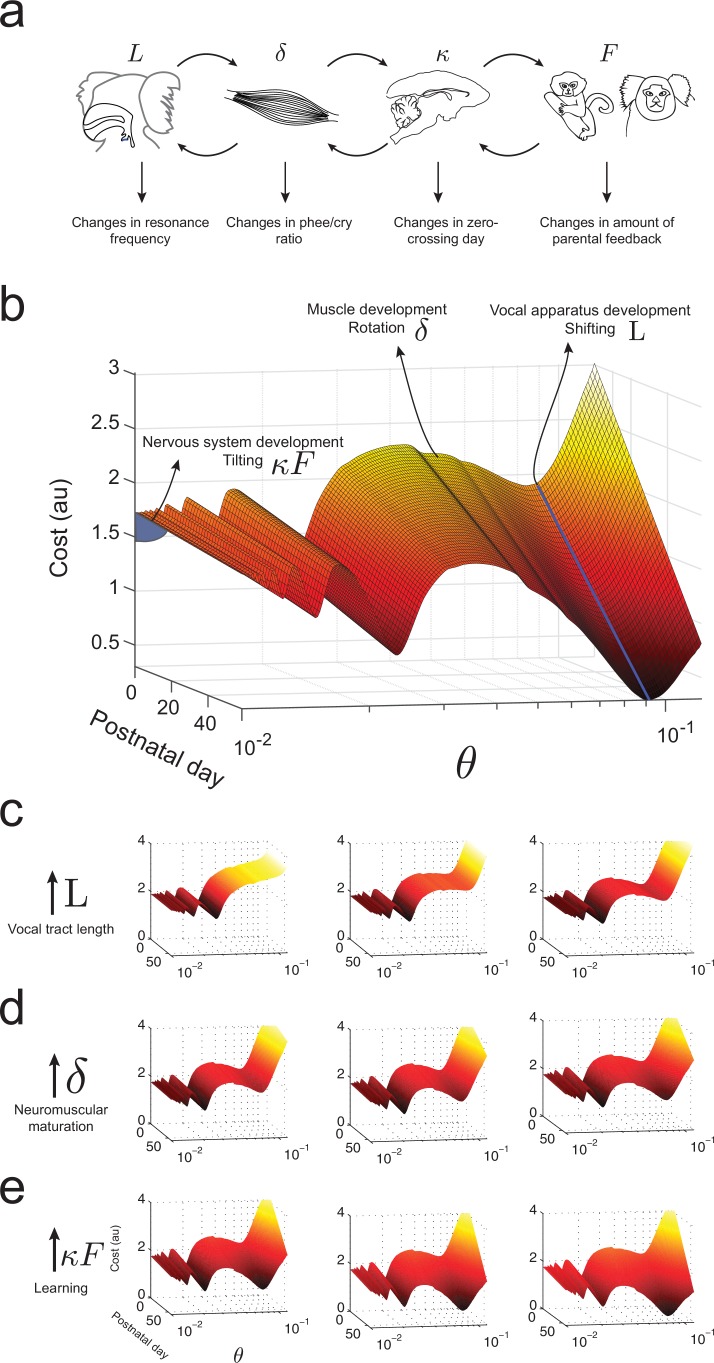
Vocal development is the adaptive coordination of the vocal apparatus, muscles, the nervous system, and social interaction. Here, we use a quantitative framework based on optimal control theory and Waddington's landscape metaphor to provide an integrated view of this process. With a biomechanical model of the marmoset monkey vocal apparatus and behavioral developmental data, we show that only the combination of the developing vocal tract, vocal apparatus muscles and nervous system can fully account for the patterns of vocal development. Together, these elements influence the shape of the monkeys' vocal developmental landscape, tilting, rotating or shifting it in different ways. We can thus use this framework to make quantitative predictions regarding how interfering factors or experimental perturbations can change the landscape within a species, or to explain comparative differences in vocal development across species.
Keywords: developmental systems; epigenetic landscape; marmoset monkey; neuromechanics; neuroscience; songbird; vocal tract resonance.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
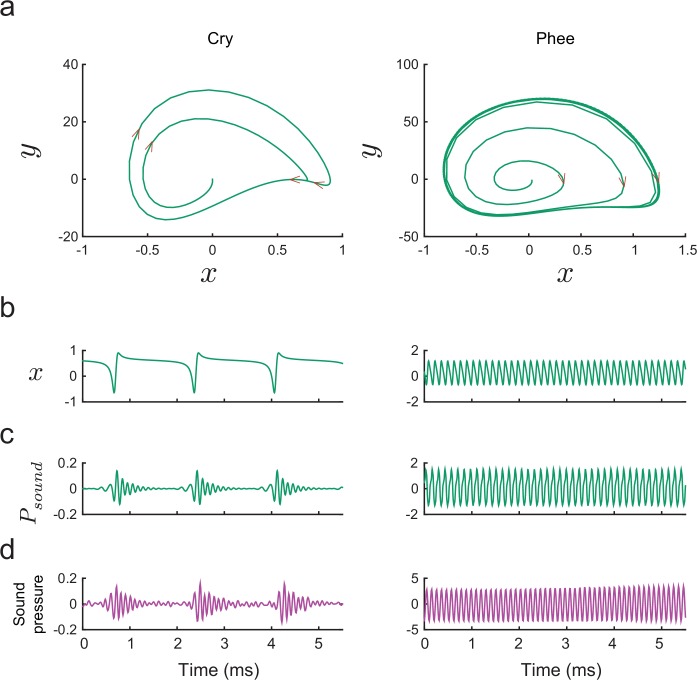
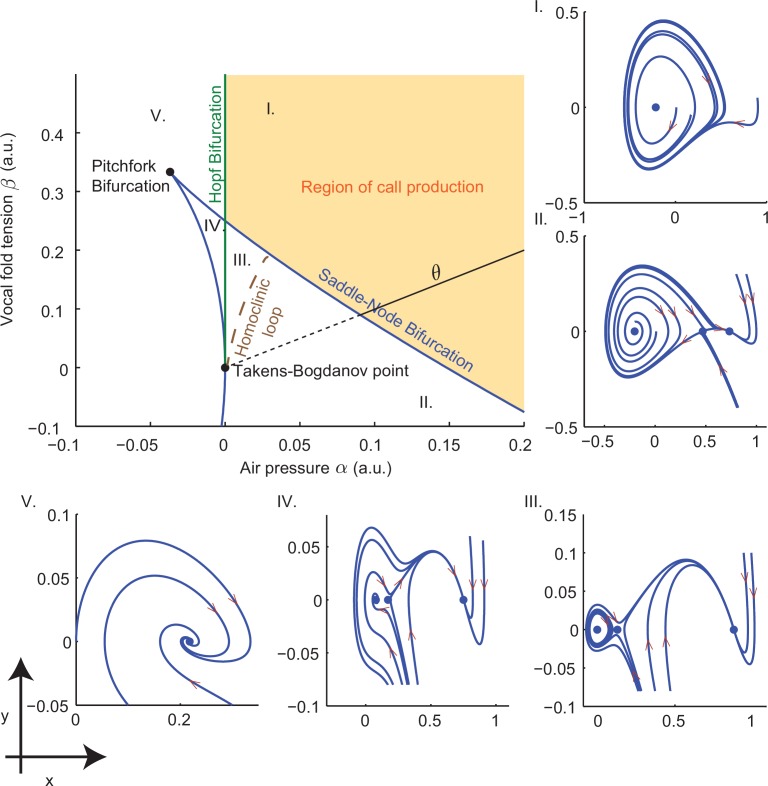
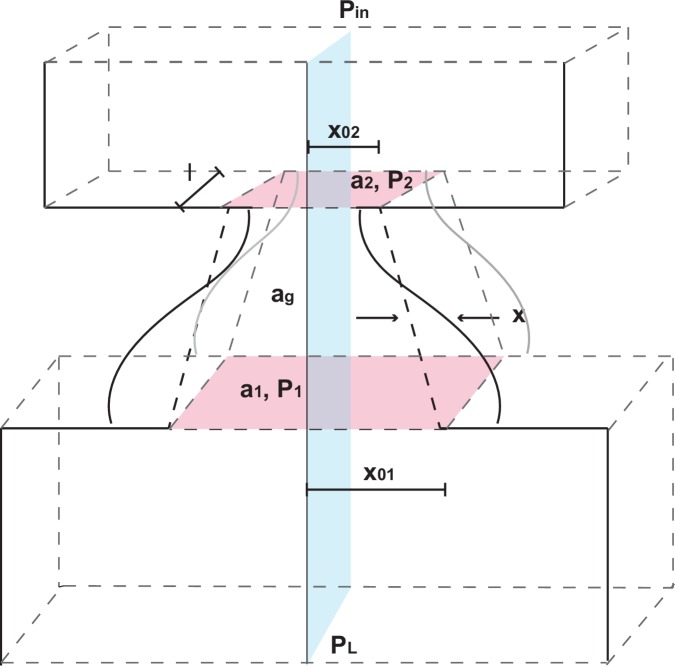
References
-
- Amador A. PhD thesis. University of Buenos Aires; 2009. Nonlinear effects in the generation of birdsong.
-
- Bezerra BM, Souto A. Structure and usage of the vocal repertoire of callithrix jacchus. International Journal of Primatology. 2008;29:671–701. doi: 10.1007/s10764-008-9250-0. - DOI
-
- Bird R, Stewart W, Lightfoot E. Transport phenomena. New York: John Wiley & Sons; 2007.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
