

Multiple shared mechanisms for homeostatic plasticity in rodent somatosensory and visual cortex
- PMID: 28093551
- PMCID: PMC5247589
- DOI: 10.1098/rstb.2016.0157
Multiple shared mechanisms for homeostatic plasticity in rodent somatosensory and visual cortex
Abstract
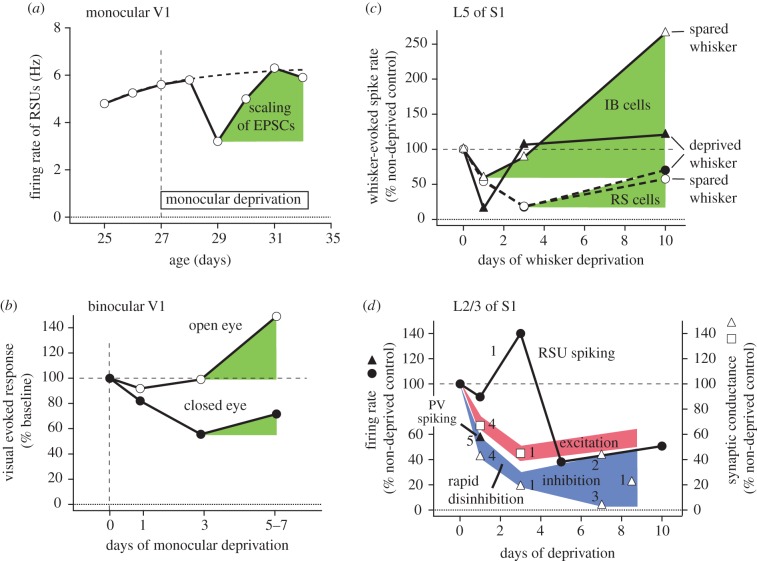
We compare the circuit and cellular mechanisms for homeostatic plasticity that have been discovered in rodent somatosensory (S1) and visual (V1) cortex. Both areas use similar mechanisms to restore mean firing rate after sensory deprivation. Two time scales of homeostasis are evident, with distinct mechanisms. Slow homeostasis occurs over several days, and is mediated by homeostatic synaptic scaling in excitatory networks and, in some cases, homeostatic adjustment of pyramidal cell intrinsic excitability. Fast homeostasis occurs within less than 1 day, and is mediated by rapid disinhibition, implemented by activity-dependent plasticity in parvalbumin interneuron circuits. These processes interact with Hebbian synaptic plasticity to maintain cortical firing rates during learned adjustments in sensory representations.This article is part of the themed issue 'Integrating Hebbian and homeostatic plasticity'.
Keywords: firing rate homeostasis; homeostatic plasticity; inhibition; sensory cortex; somatosensory cortex; whisker.
© 2017 The Author(s).
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
