

Predictive Markers Guide Differentiation to Improve Graft Outcome in Clinical Translation of hESC-Based Therapy for Parkinson's Disease
- PMID: 28094017
- PMCID: PMC5222722
- DOI: 10.1016/j.stem.2016.09.004
Predictive Markers Guide Differentiation to Improve Graft Outcome in Clinical Translation of hESC-Based Therapy for Parkinson's Disease
Abstract
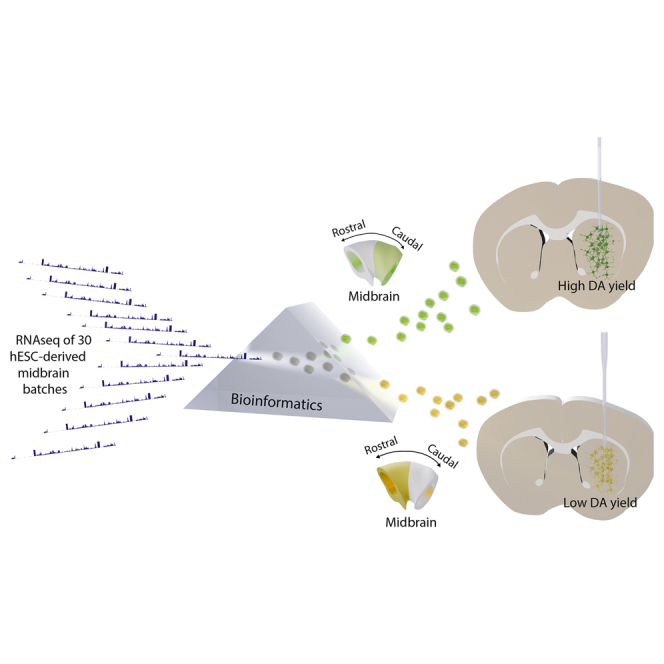
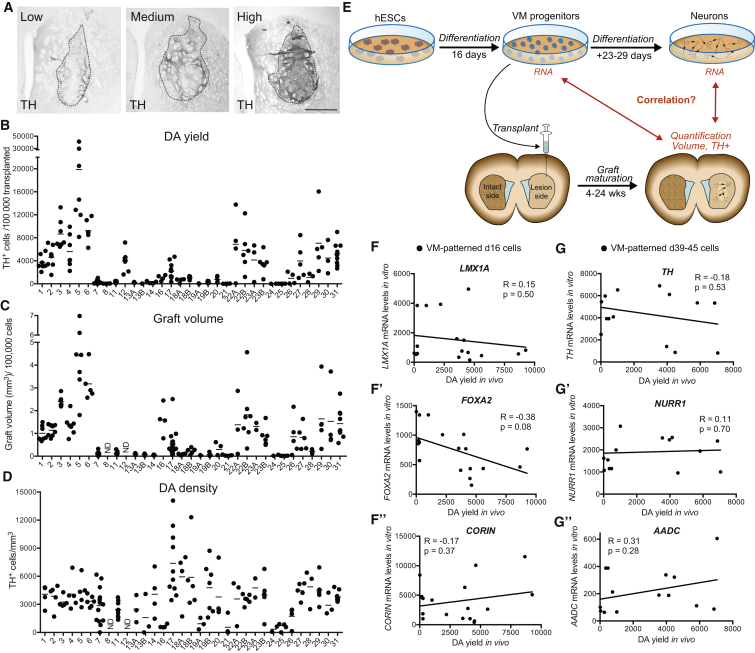
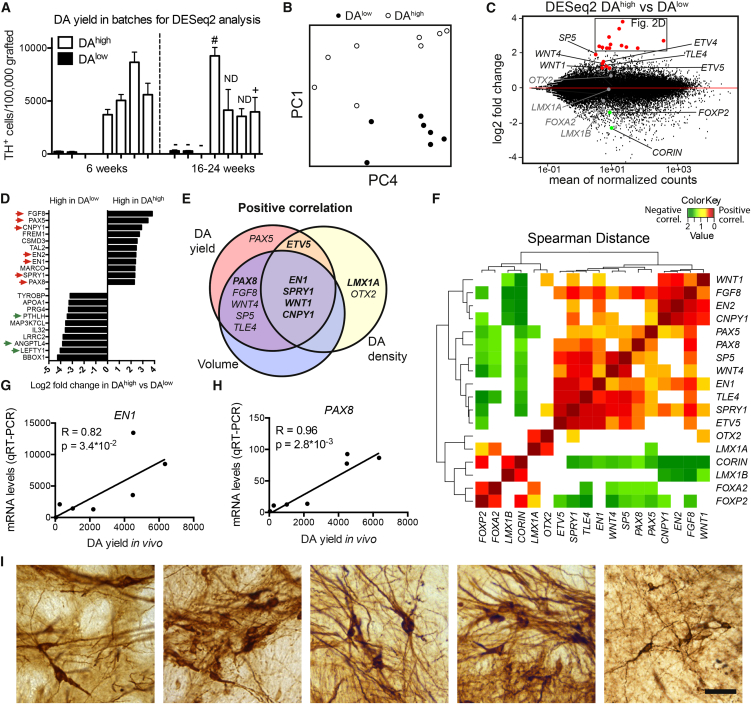
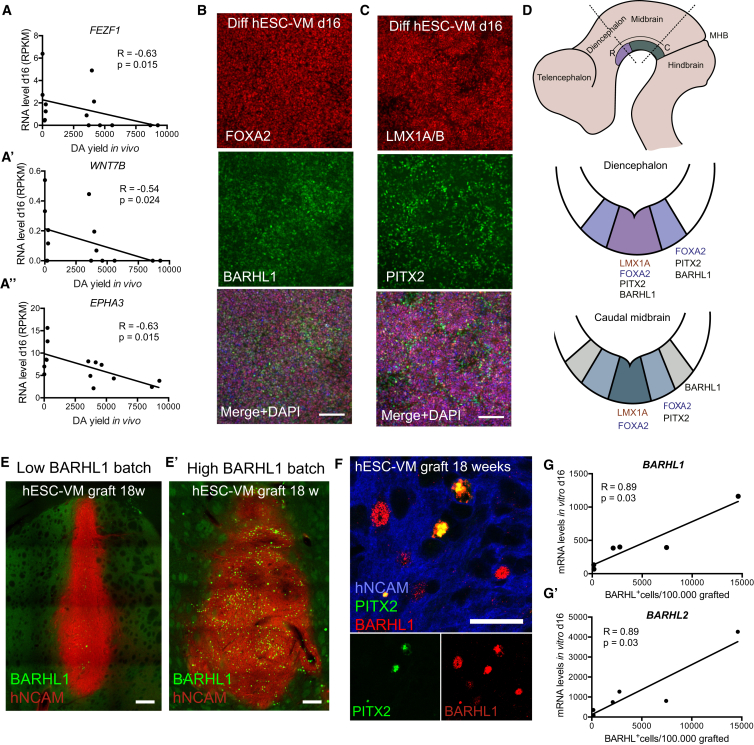
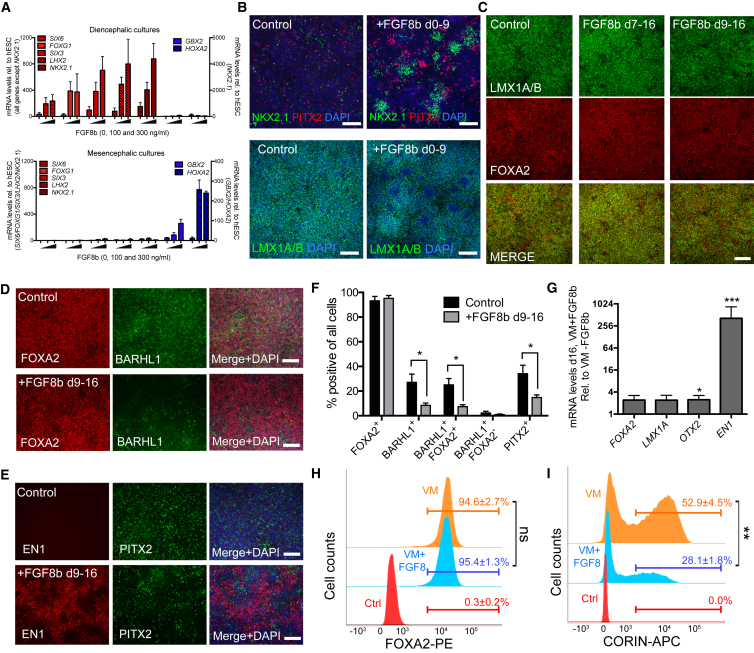
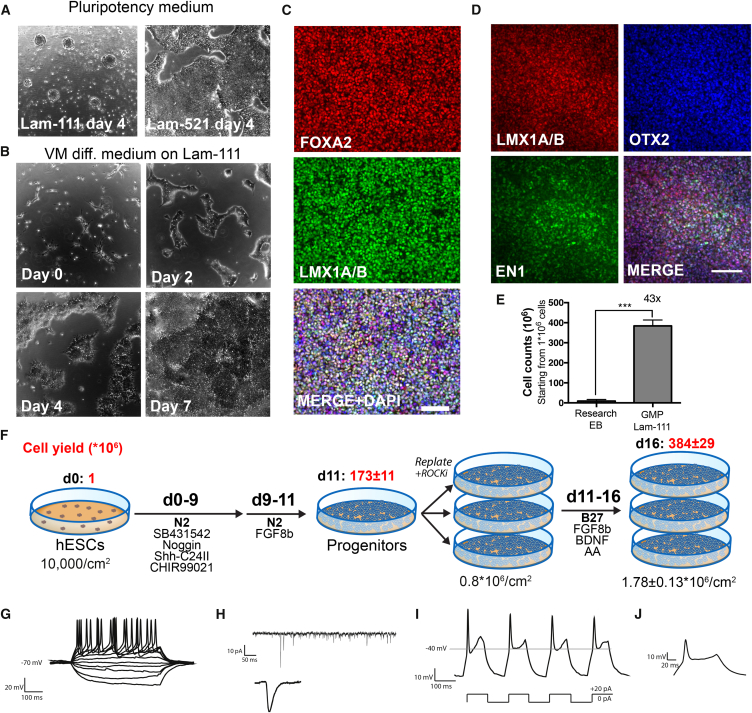
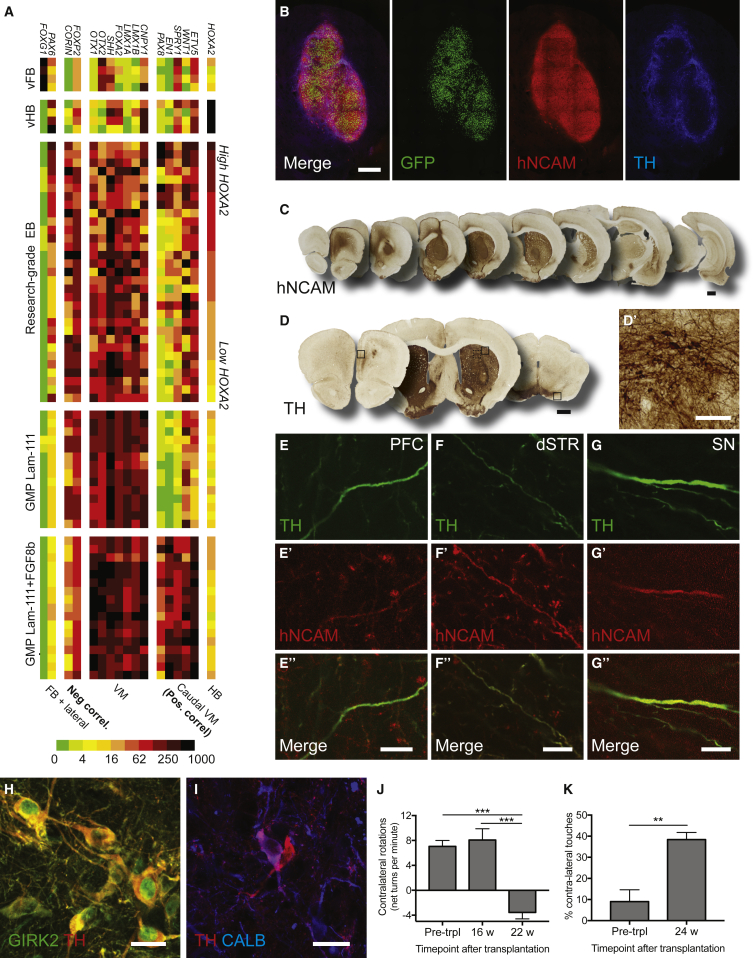
Stem cell treatments for neurodegenerative diseases are expected to reach clinical trials soon. Most of the approaches currently under development involve transplantation of immature progenitors that subsequently undergo phenotypic and functional maturation in vivo, and predicting the long-term graft outcome already at the progenitor stage remains a challenge. Here, we took an unbiased approach to identify predictive markers expressed in dopamine neuron progenitors that correlate with graft outcome in an animal model of Parkinson's disease through gene expression analysis of >30 batches of grafted human embryonic stem cell (hESC)-derived progenitors. We found that many of the commonly used markers did not accurately predict in vivo subtype-specific maturation. Instead, we identified a specific set of markers associated with the caudal midbrain that correlate with high dopaminergic yield after transplantation in vivo. Using these markers, we developed a good manufacturing practice (GMP) differentiation protocol for highly efficient and reproducible production of transplantable dopamine progenitors from hESCs.
Keywords: FGF8; GMP; MHB; Parkinson’s disease; STN; VM; diencephalon; dopaminergic neurons; good manufacturing practice; hESCs; human embryonic stem cells; midbrain-hindbrain boundary; subthalamic nucleus; transplantation; ventral midbrain.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
Seq-ing Markers of Midbrain Dopamine Neurons.Cell Stem Cell. 2017 Jan 5;20(1):11-12. doi: 10.1016/j.stem.2016.12.014. Cell Stem Cell. 2017. PMID: 28061349
References
-
- Arenas E., Denham M., Villaescusa J.C. How to make a midbrain dopaminergic neuron. Development. 2015;142:1918–1936. - PubMed
-
- Barker R.A., Drouin-Ouellet J., Parmar M. Cell-based therapies for Parkinson disease—past insights and future potential. Nat. Rev. Neurol. 2015;11:492–503. - PubMed
-
- Chakradhar S. An eye to the future: researchers debate best path for stem cell-derived therapies. Nat. Med. 2016;22:116–119. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
