

Centriole Remodeling during Spermiogenesis in Drosophila
- PMID: 28094036
- PMCID: PMC5245371
- DOI: 10.1016/j.cub.2016.07.006
Centriole Remodeling during Spermiogenesis in Drosophila
Abstract
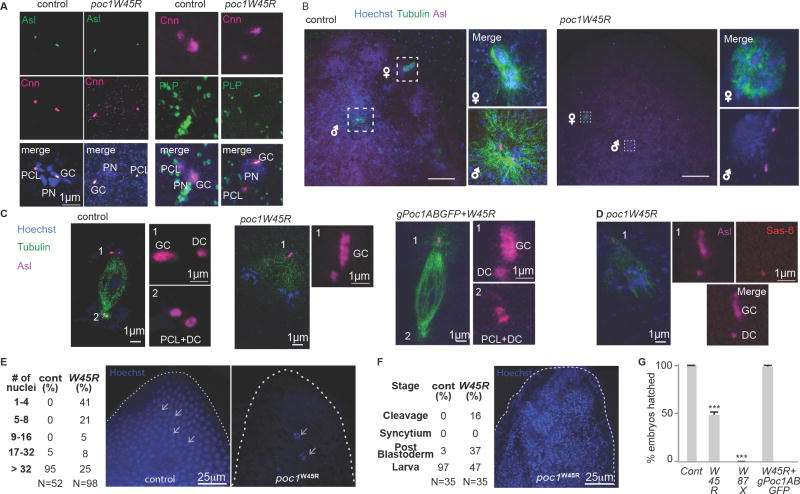
The first cell of an animal (zygote) requires centrosomes that are assembled from paternally inherited centrioles and maternally inherited pericentriolar material (PCM) [1]. In some animals, sperm centrioles with typical ultrastructure are the origin of the first centrosomes in the zygote [2-4]. In other animals, however, sperm centrioles lose their proteins and are thought to be degenerated and non-functional during spermiogenesis [5, 6]. Here, we show that the two sperm centrioles (the giant centriole [GC] and the proximal centriole-like structure [PCL]) in Drosophila melanogaster are remodeled during spermiogenesis through protein enrichment and ultrastructure modification in parallel to previously described centrosomal reduction [7]. We found that the ultrastructure of the matured sperm (spermatozoa) centrioles is modified dramatically and that the PCL does not resemble a typical centriole. We also describe a new phenomenon of Poc1 enrichment of the atypical centrioles in the spermatozoa. Using various mutants, protein expression during spermiogenesis, and RNAi knockdown of paternal Poc1, we found that paternal Poc1 enrichment is essential for the formation of centrioles during spermiogenesis and for the formation of centrosomes after fertilization in the zygote. Altogether, these findings demonstrate that the sperm centrioles are remodeled both in their protein composition and in ultrastructure, yet they are functional and are essential for normal embryogenesis in Drosophila.
Keywords: PCL; Poc1; centriole; centrosome; centrosome reaction; embryo; fertilization; giant centriole; sperm; zygote.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures



References
-
- Terada Y, Schatten G, Hasegawa H, Yaegashi N. Essential roles of the sperm centrosome in human fertilization: developing the therapy for fertilization failure due to sperm centrosomal dysfunction. Tohoku J Exp Med. 2010;220:247–258. - PubMed
-
- O’Connell KF, Caron C, Kopish KR, Hurd DD, Kemphues KJ, Li Y, White JG. The C. elegans zyg-1 gene encodes a regulator of centrosome duplication with distinct maternal and paternal roles in the embryo. Cell. 2001;105:547–558. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
