

Common developmental genome deprogramming in schizophrenia - Role of Integrative Nuclear FGFR1 Signaling (INFS)
- PMID: 28094170
- PMCID: PMC5507209
- DOI: 10.1016/j.schres.2016.12.012
Common developmental genome deprogramming in schizophrenia - Role of Integrative Nuclear FGFR1 Signaling (INFS)
Abstract
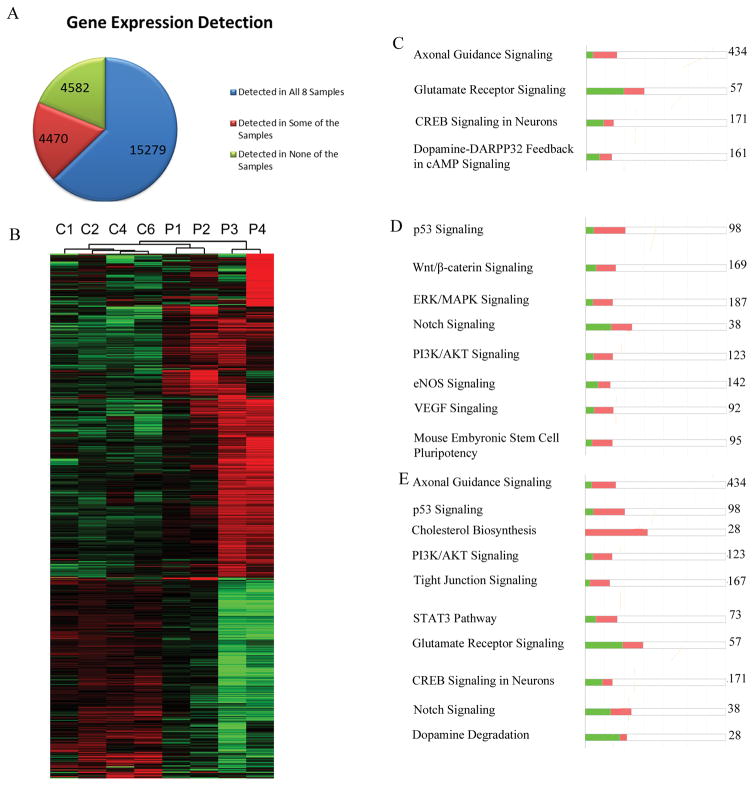
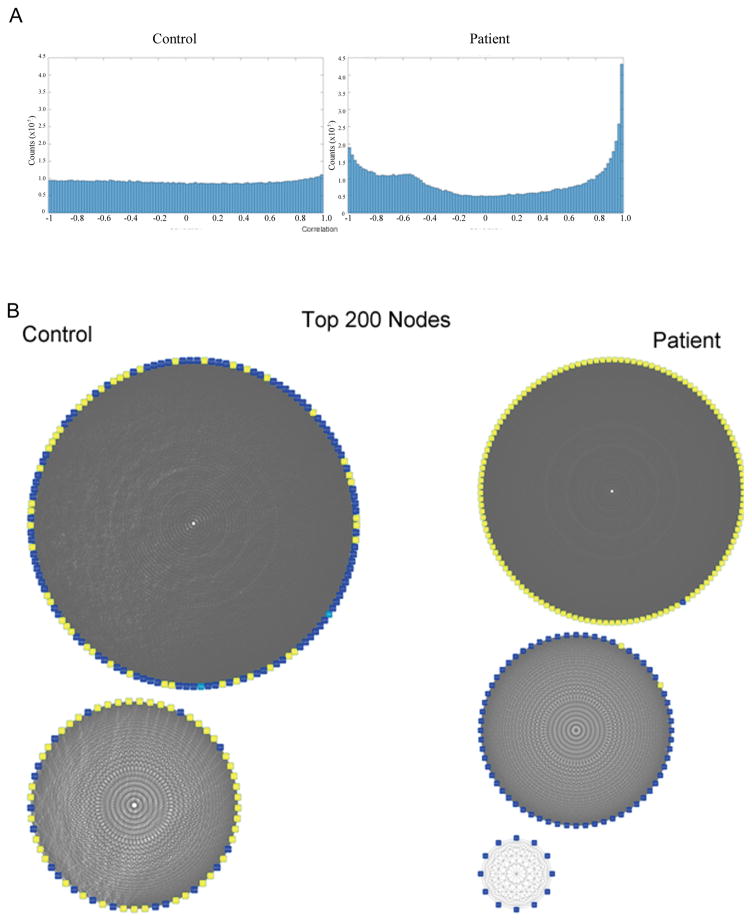
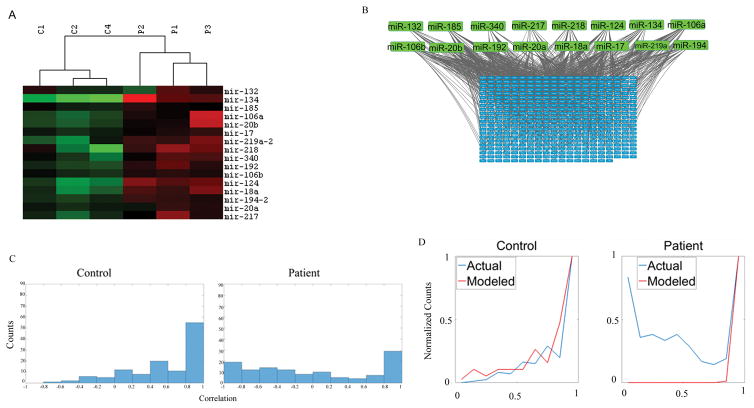
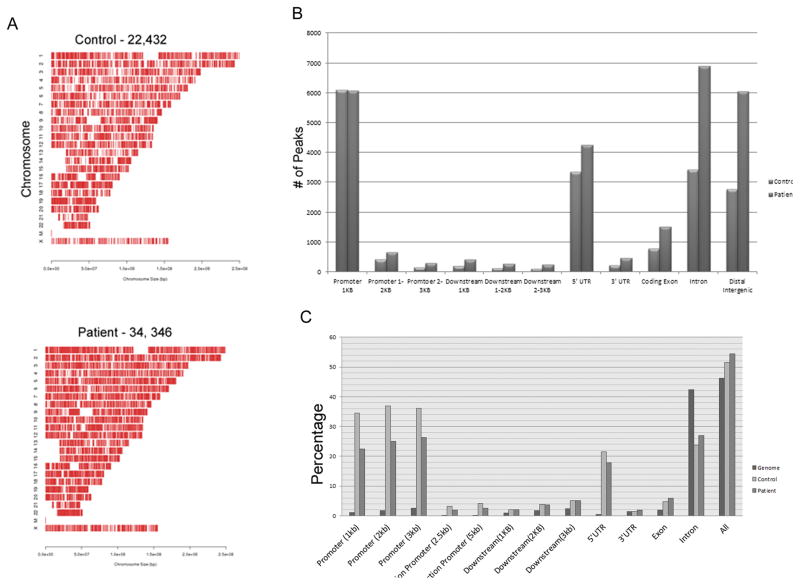
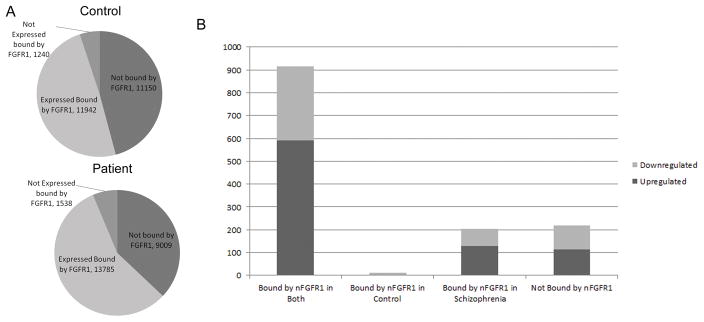
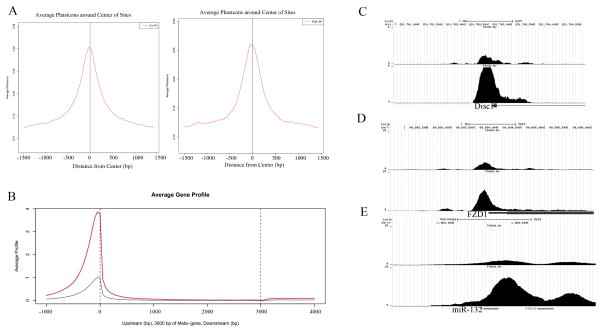
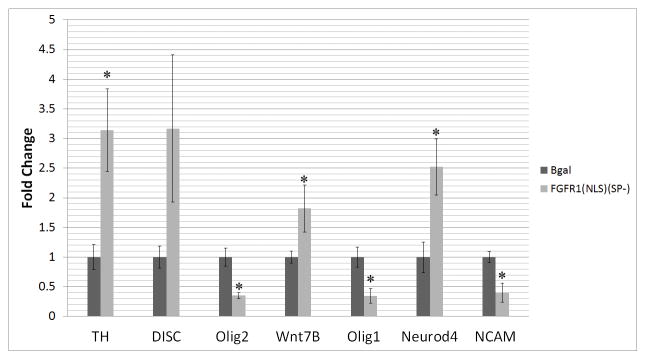
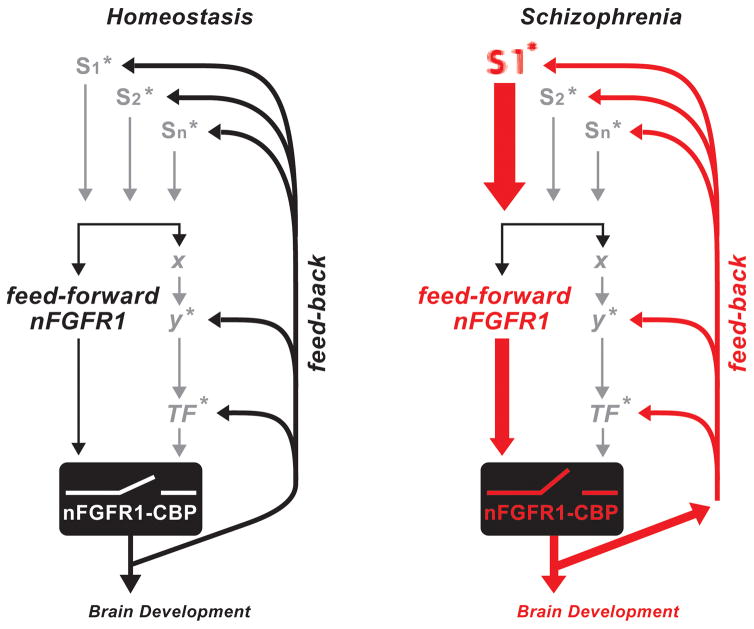
The watershed-hypothesis of schizophrenia asserts that over 200 different mutations dysregulate distinct pathways that converge on an unspecified common mechanism(s) that controls disease ontogeny. Consistent with this hypothesis, our RNA-sequencing of neuron committed cells (NCCs) differentiated from established iPSCs of 4 schizophrenia patients and 4 control subjects uncovered a dysregulated transcriptome of 1349 mRNAs common to all patients. Data reveals a global dysregulation of developmental genome, deconstruction of coordinated mRNA networks, and the formation of aberrant, new coordinated mRNA networks indicating a concerted action of the responsible factor(s). Sequencing of miRNA transcriptomes demonstrated an overexpression of 16 miRNAs and deconstruction of interactive miRNA-mRNA networks in schizophrenia NCCs. ChiPseq revealed that the nuclear (n) form of FGFR1, a pan-ontogenic regulator, is overexpressed in schizophrenia NCCs and overtargets dysregulated mRNA and miRNA genes. The nFGFR1 targeted 54% of all human gene promoters and 84.4% of schizophrenia dysregulated genes. The upregulated genes reside within major developmental pathways that control neurogenesis and neuron formation, whereas downregulated genes are involved in oligodendrogenesis. Our results indicate (i) an early (preneuronal) genomic etiology of schizophrenia, (ii) dysregulated genes and new coordinated gene networks are common to unrelated cases of schizophrenia, (iii) gene dysregulations are accompanied by increased nFGFR1-genome interactions, and (iv) modeling of increased nFGFR1 by an overexpression of a nFGFR1 lead to up or downregulation of selected genes as observed in schizophrenia NCCs. Together our results designate nFGFR1 signaling as a potential common dysregulated mechanism in investigated patients and potential therapeutic target in schizophrenia.
Keywords: Developmental hypothesis; Fibroblast Growth Factor Receptor 1; Genome regulation; Schizophrenia; miRNA.
Copyright © 2016 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Figures
References
-
- Akbarian S, Bunney WE, Jr, Potkin SG, Wigal SB, Hagman JO, Sandman CA, Jones EG. Altered distribution of nicotinamide-adenine dinucleotide phosphate-diaphorase cells in frontal lobe of schizophrenics implies disturbances of cortical development. Archives of general psychiatry. 1993;50(3):169–177. - PubMed
-
- Arnold SE, Ruscheinsky DD, Han LY. Further evidence of abnormal cytoarchitecture of the entorhinal cortex in schizophrenia using spatial point pattern analyses. Biological psychiatry. 1997;42(8):639–647. - PubMed
-
- Baron O, Forthmann B, Lee YW, Terranova C, Ratzka A, Stachowiak EK, Grothe C, Claus P, Stachowiak MK. Cooperation of nuclear fibroblast growth factor receptor 1 and nurr1 offers new interactive mechanism in postmitotic development of mesencephalic dopaminergic neurons. The Journal of biological chemistry. 2012b;287(24):19827–19840. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
