

Influences of Host Community Characteristics on Borrelia burgdorferi Infection Prevalence in Blacklegged Ticks
- PMID: 28095423
- PMCID: PMC5241014
- DOI: 10.1371/journal.pone.0167810
Influences of Host Community Characteristics on Borrelia burgdorferi Infection Prevalence in Blacklegged Ticks
Abstract
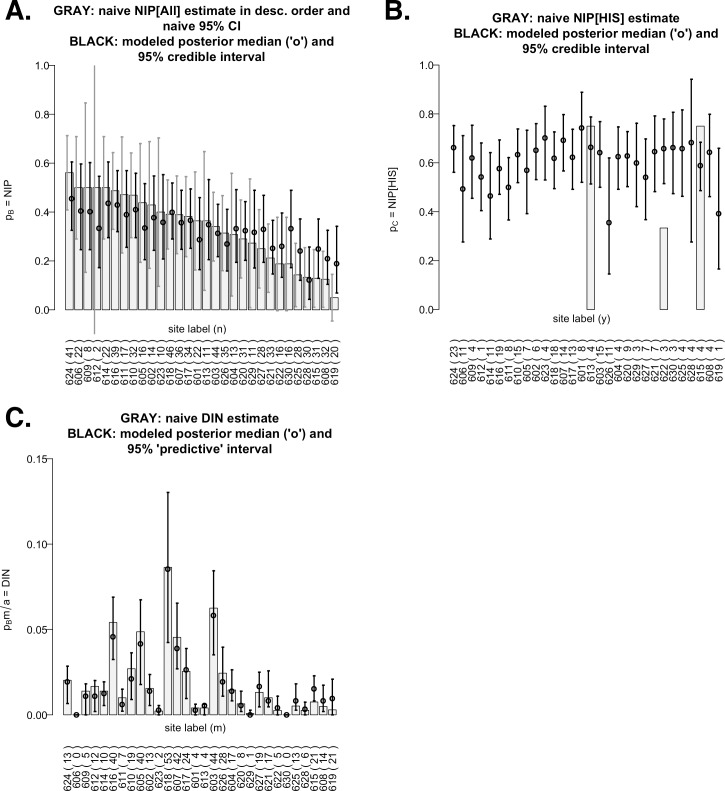
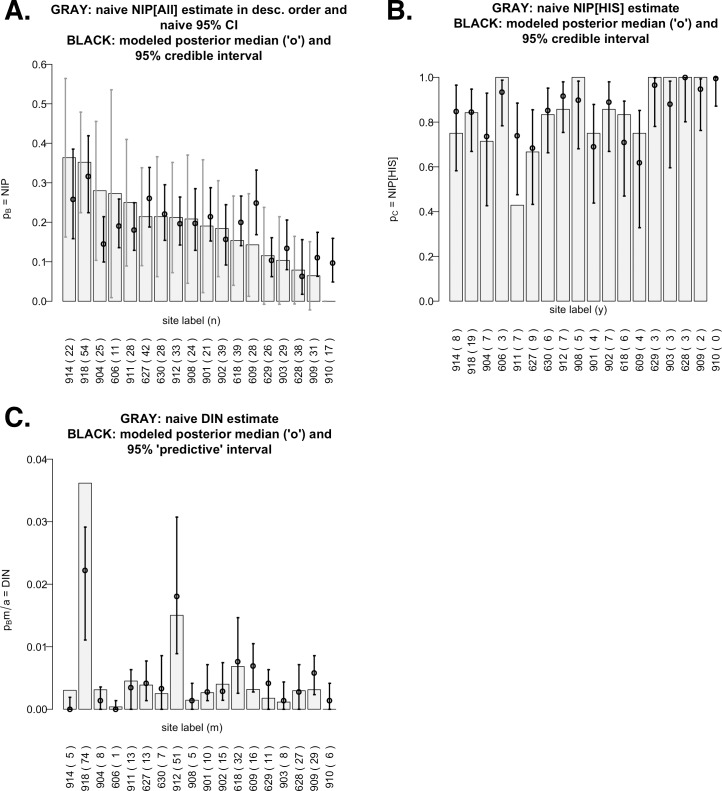
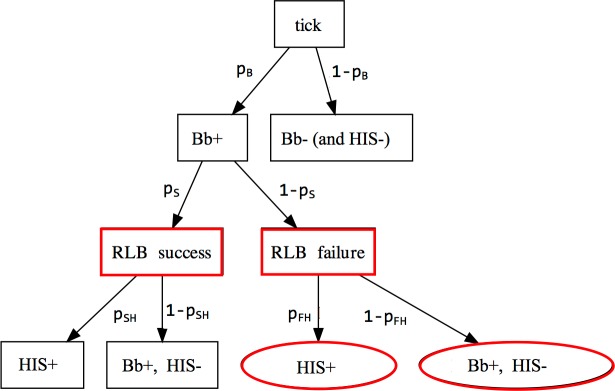
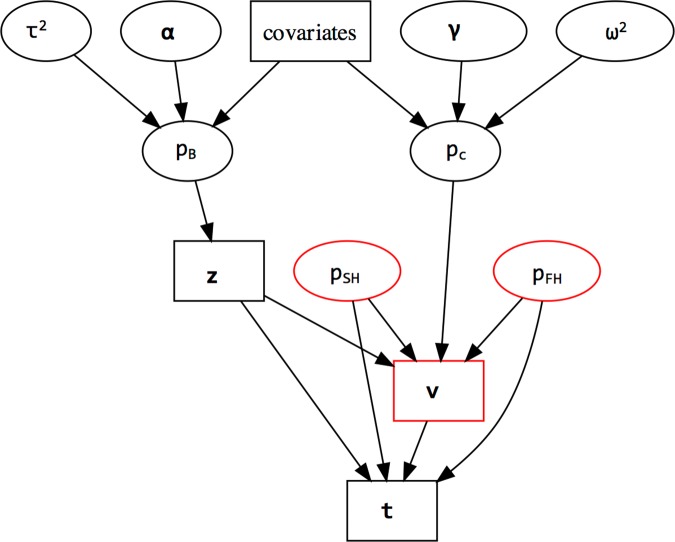
Lyme disease is a major vector-borne bacterial disease in the USA. The disease is caused by Borrelia burgdorferi, and transmitted among hosts and humans, primarily by blacklegged ticks (Ixodes scapularis). The ~25 B. burgdorferi genotypes, based on genotypic variation of their outer surface protein C (ospC), can be phenotypically separated as strains that primarily cause human diseases-human invasive strains (HIS)-or those that rarely do. Additionally, the genotypes are non-randomly associated with host species. The goal of this study was to examine the extent to which phenotypic outcomes of B. burgdorferi could be explained by the host communities fed upon by blacklegged ticks. In 2006 and 2009, we determined the host community composition based on abundance estimates of the vertebrate hosts, and collected host-seeking nymphal ticks in 2007 and 2010 to determine the ospC genotypes within infected ticks. We regressed instances of B. burgdorferi phenotypes on site-specific characteristics of host communities by constructing Bayesian hierarchical models that properly handled missing data. The models provided quantitative support for the relevance of host composition on Lyme disease risk pertaining to B. burgdorferi prevalence (i.e. overall nymphal infection prevalence, or NIPAll) and HIS prevalence among the infected ticks (NIPHIS). In each year, NIPAll and NIPHIS was found to be associated with host relative abundances and diversity. For mice and chipmunks, the association with NIPAll was positive, but tended to be negative with NIPHIS in both years. However, the direction of association between shrew relative abundance with NIPAll or NIPHIS differed across the two years. And, diversity (H') had a negative association with NIPAll, but positive association with NIPHIS in both years. Our analyses highlight that the relationships between the relative abundances of three primary hosts and the community diversity with NIPAll, and NIPHIS, are variable in time and space, and that disease risk inference, based on the role of host community, changes when we examine risk overall or at the phenotypic level. Our discussion focuses on the observed relationships between prevalence and host community characteristics and how they substantiate the ecological understanding of phenotypic Lyme disease risk.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
