

Cartilage to bone transformation during fracture healing is coordinated by the invading vasculature and induction of the core pluripotency genes
- PMID: 28096214
- PMCID: PMC5394763
- DOI: 10.1242/dev.130807
Cartilage to bone transformation during fracture healing is coordinated by the invading vasculature and induction of the core pluripotency genes
Abstract
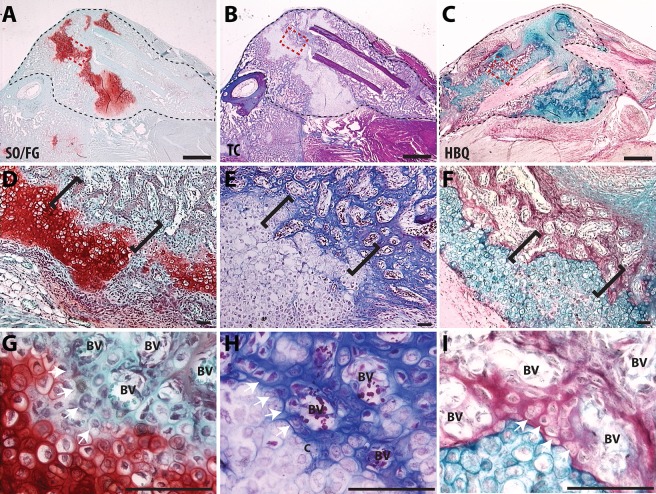
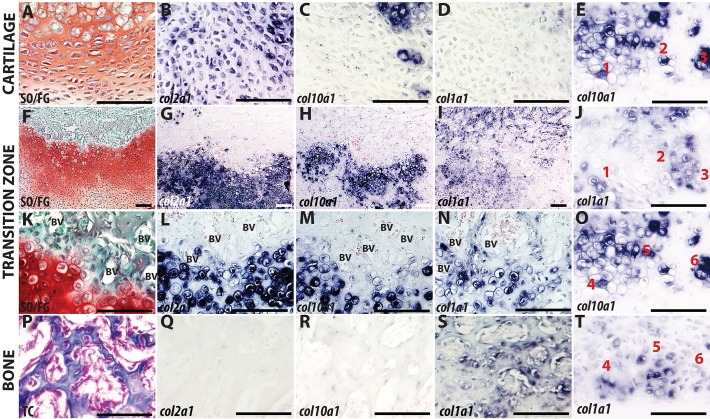
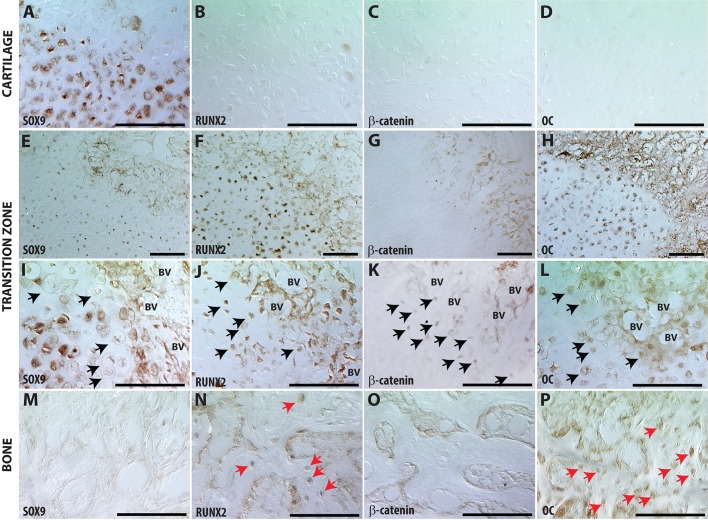
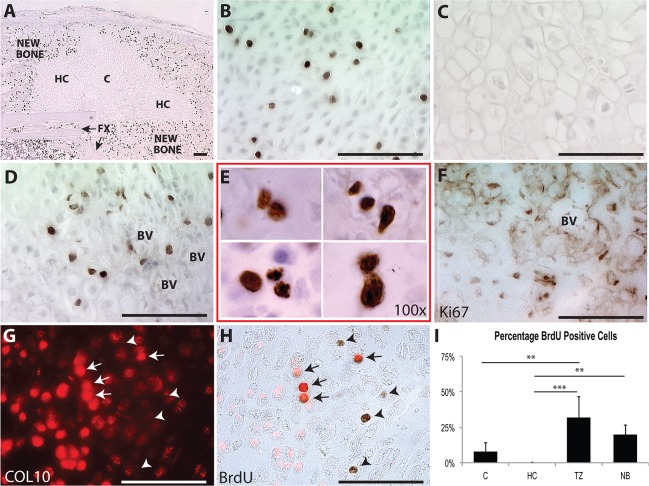
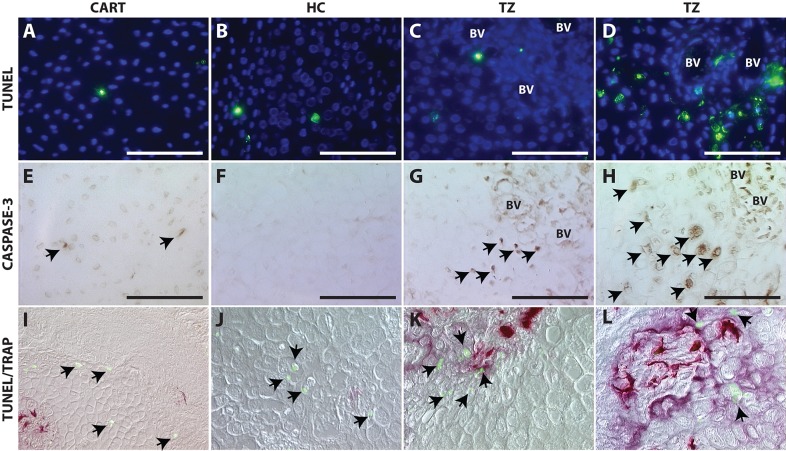
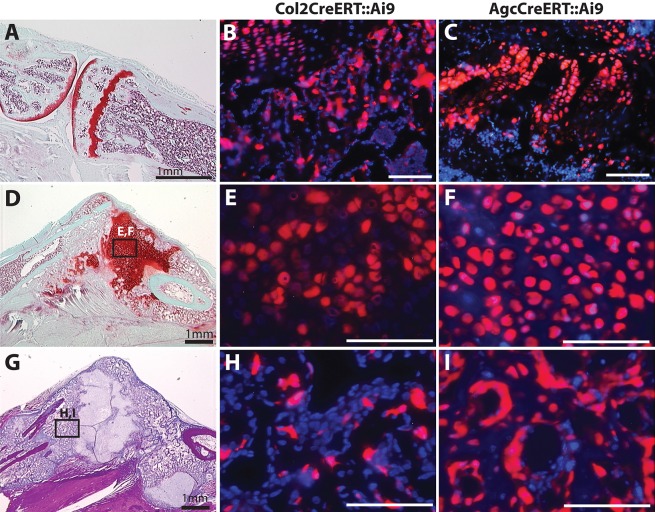
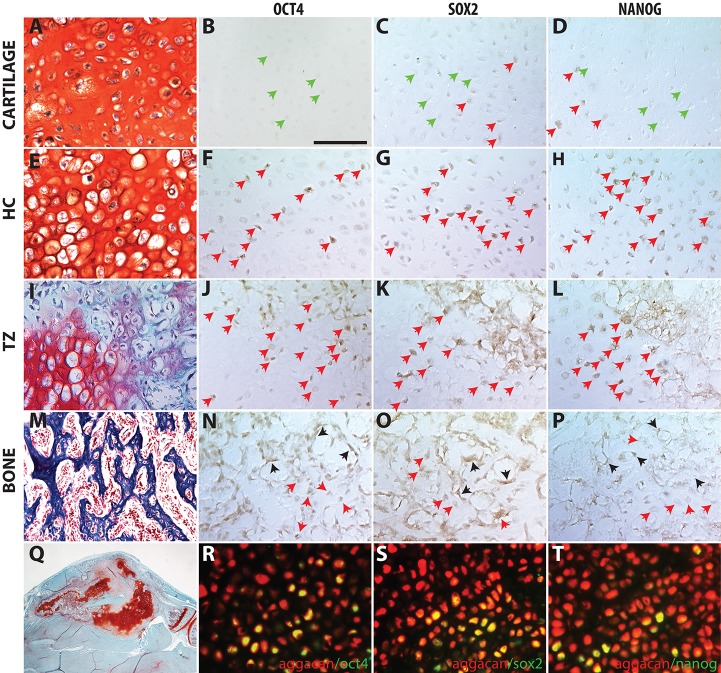
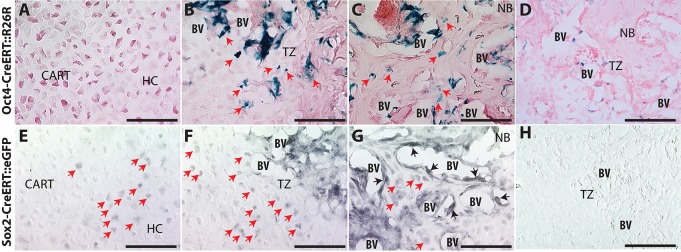
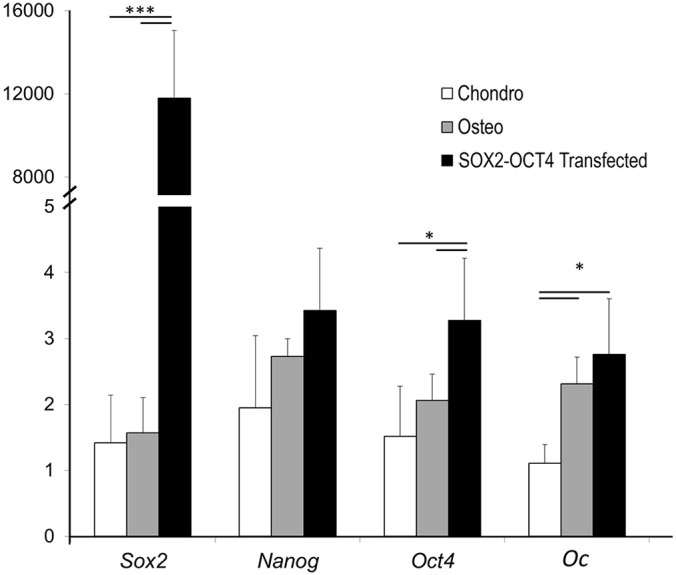
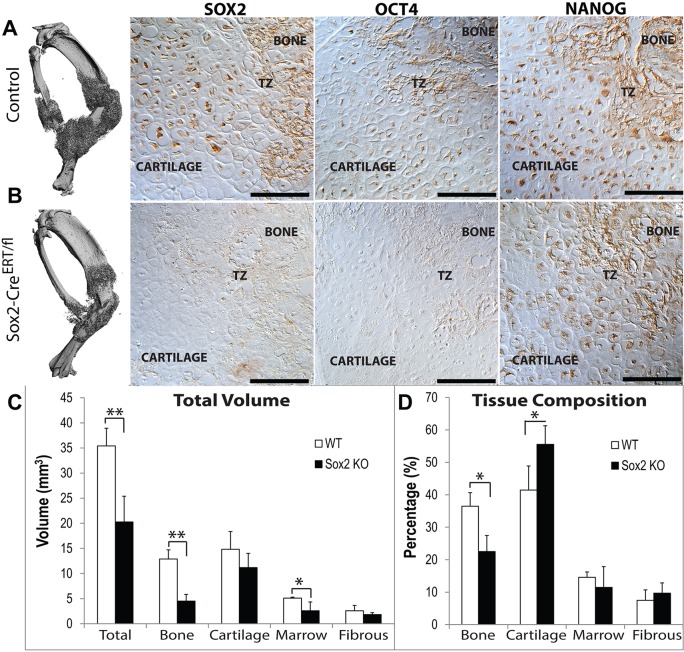
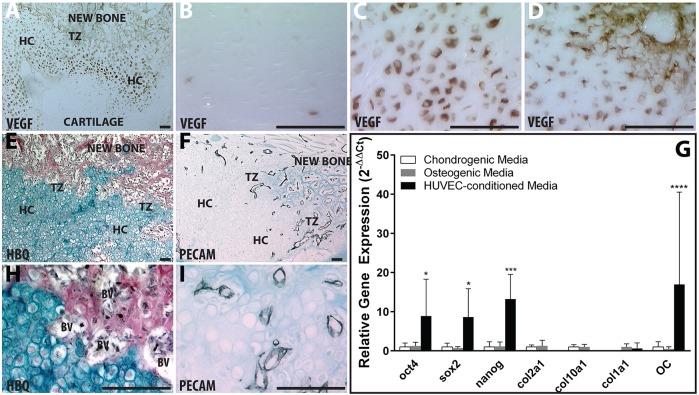
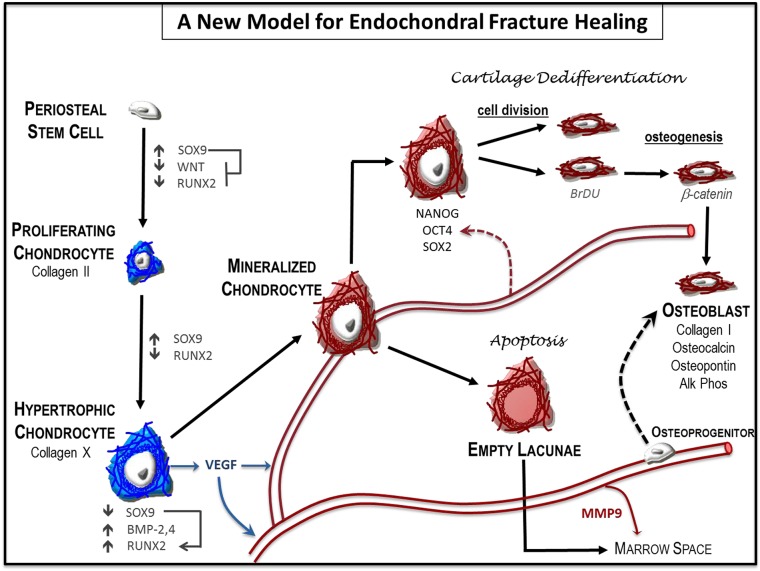
Fractures heal predominantly through the process of endochondral ossification. The classic model of endochondral ossification holds that chondrocytes mature to hypertrophy, undergo apoptosis and new bone forms by invading osteoprogenitors. However, recent data demonstrate that chondrocytes transdifferentiate to osteoblasts in the growth plate and during regeneration, yet the mechanism(s) regulating this process remain unknown. Here, we show a spatially-dependent phenotypic overlap between hypertrophic chondrocytes and osteoblasts at the chondro-osseous border in the fracture callus, in a region we define as the transition zone (TZ). Hypertrophic chondrocytes in the TZ activate expression of the pluripotency factors [Sox2, Oct4 (Pou5f1), Nanog], and conditional knock-out of Sox2 during fracture healing results in reduction of the fracture callus and a delay in conversion of cartilage to bone. The signal(s) triggering expression of the pluripotency genes are unknown, but we demonstrate that endothelial cell conditioned medium upregulates these genes in ex vivo fracture cultures, supporting histological evidence that transdifferentiation occurs adjacent to the vasculature. Elucidating the cellular and molecular mechanisms underlying fracture repair is important for understanding why some fractures fail to heal and for developing novel therapeutic interventions.
Keywords: Chondrocyte transformation; Endochondral ossification; Fracture repair; Pluripotency programs.
© 2017. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
References
-
- Aspenberg P., Genant H. K., Johansson T., Nino A. J., See K., Krohn K., García-Hernández P. A., Recknor C. P., Einhorn T. A., Dalsky G. P. et al. (2010). Teriparatide for acceleration of fracture repair in humans: a prospective, randomized, double-blind study of 102 postmenopausal women with distal radial fractures. J. Bone Miner. Res. 25, 404-414. 10.1359/jbmr.090731 - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
