

Efficient CRISPR/Cas9-assisted gene targeting enables rapid and precise genetic manipulation of mammalian neural stem cells
- PMID: 28096221
- PMCID: PMC5312033
- DOI: 10.1242/dev.140855
Efficient CRISPR/Cas9-assisted gene targeting enables rapid and precise genetic manipulation of mammalian neural stem cells
Abstract
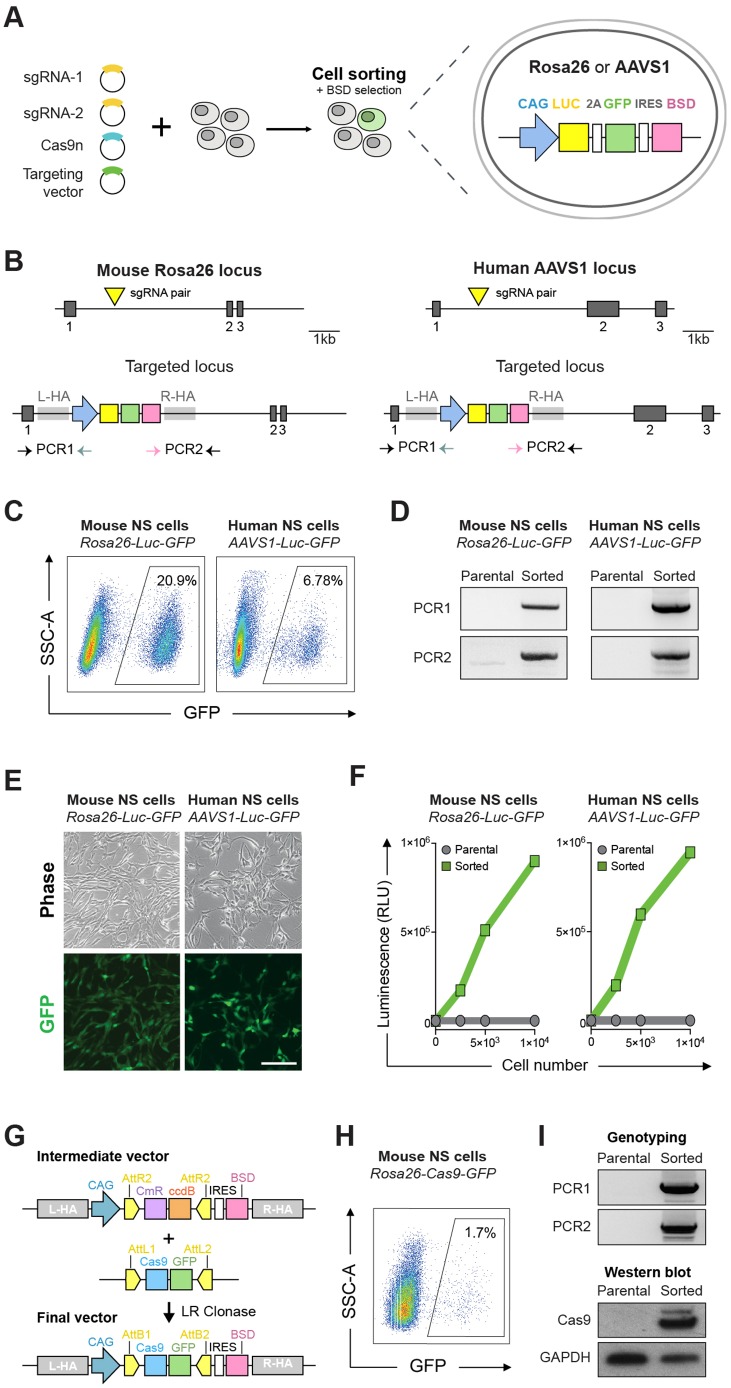
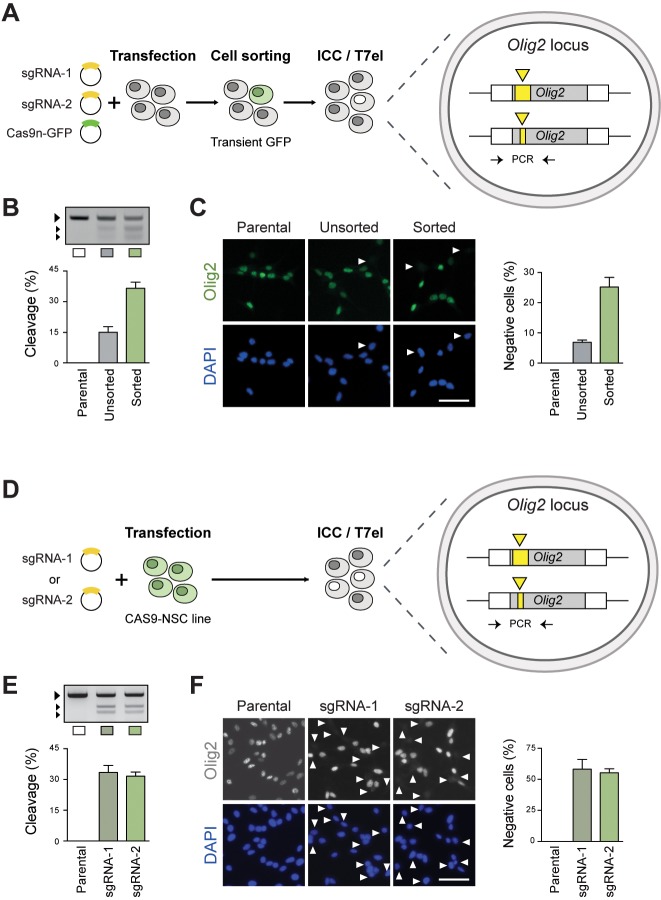
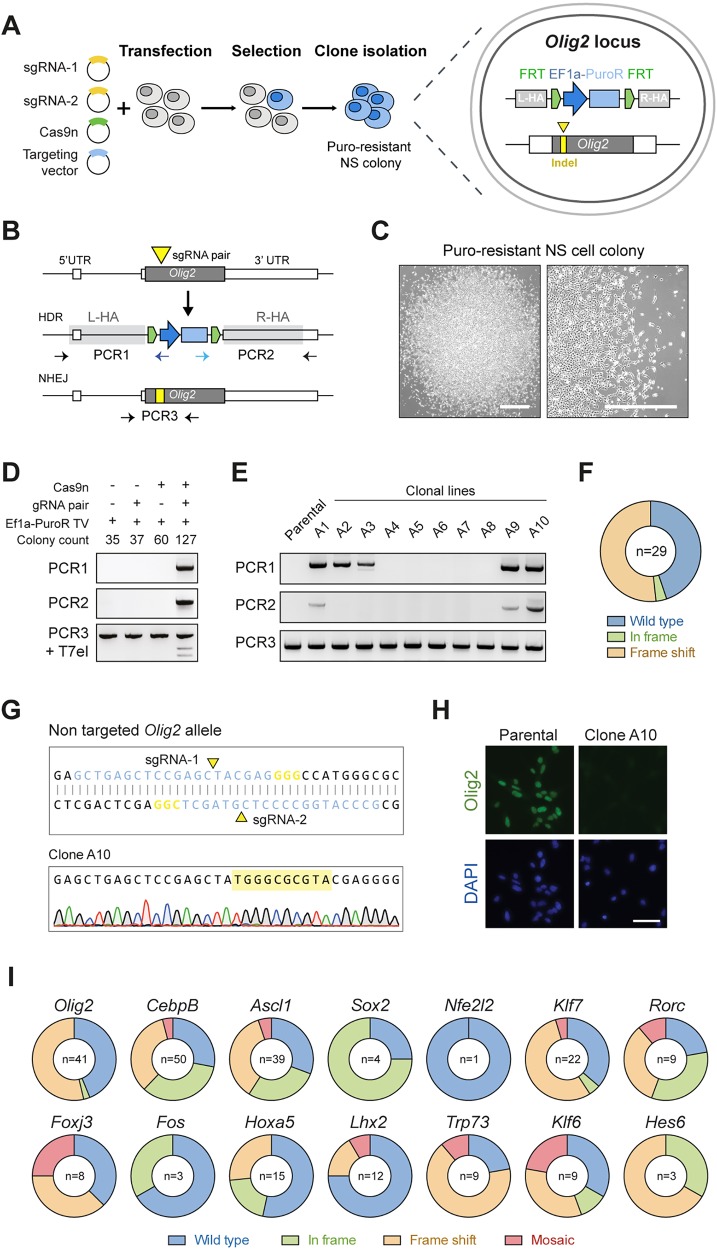
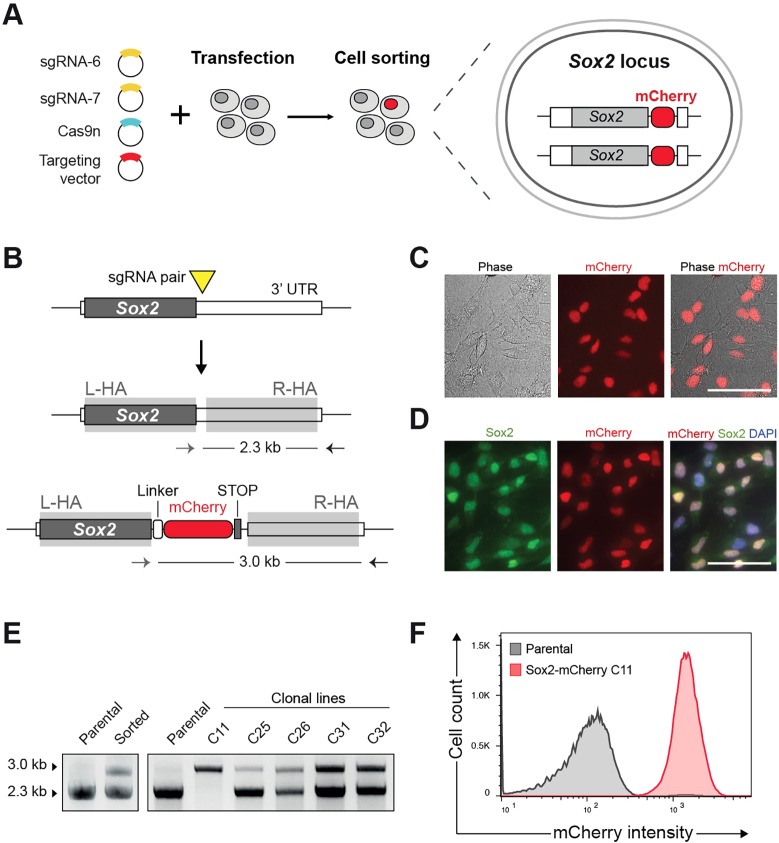
Mammalian neural stem cell (NSC) lines provide a tractable model for discovery across stem cell and developmental biology, regenerative medicine and neuroscience. They can be derived from foetal or adult germinal tissues and continuously propagated in vitro as adherent monolayers. NSCs are clonally expandable, genetically stable, and easily transfectable - experimental attributes compatible with targeted genetic manipulations. However, gene targeting, which is crucial for functional studies of embryonic stem cells, has not been exploited to date in NSC lines. Here, we deploy CRISPR/Cas9 technology to demonstrate a variety of sophisticated genetic modifications via gene targeting in both mouse and human NSC lines, including: (1) efficient targeted transgene insertion at safe harbour loci (Rosa26 and AAVS1); (2) biallelic knockout of neurodevelopmental transcription factor genes; (3) simple knock-in of epitope tags and fluorescent reporters (e.g. Sox2-V5 and Sox2-mCherry); and (4) engineering of glioma mutations (TP53 deletion; H3F3A point mutations). These resources and optimised methods enable facile and scalable genome editing in mammalian NSCs, providing significant new opportunities for functional genetic analysis.
Keywords: CRISPR/Cas9; Epitope tagging; Gene targeting; Genome editing; Glioblastoma; Homologous recombination; Neural stem cell; Transcription factor.
© 2017. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
