

A natural product inhibits the initiation of α-synuclein aggregation and suppresses its toxicity
- PMID: 28096355
- PMCID: PMC5307473
- DOI: 10.1073/pnas.1610586114
A natural product inhibits the initiation of α-synuclein aggregation and suppresses its toxicity
Erratum in
-
Correction for Perni et al., A natural product inhibits the initiation of α-synuclein aggregation and suppresses its toxicity.Proc Natl Acad Sci U S A. 2017 Mar 21;114(12):E2543. doi: 10.1073/pnas.1701964114. Epub 2017 Mar 6. Proc Natl Acad Sci U S A. 2017. PMID: 28265088 Free PMC article. No abstract available.
Abstract
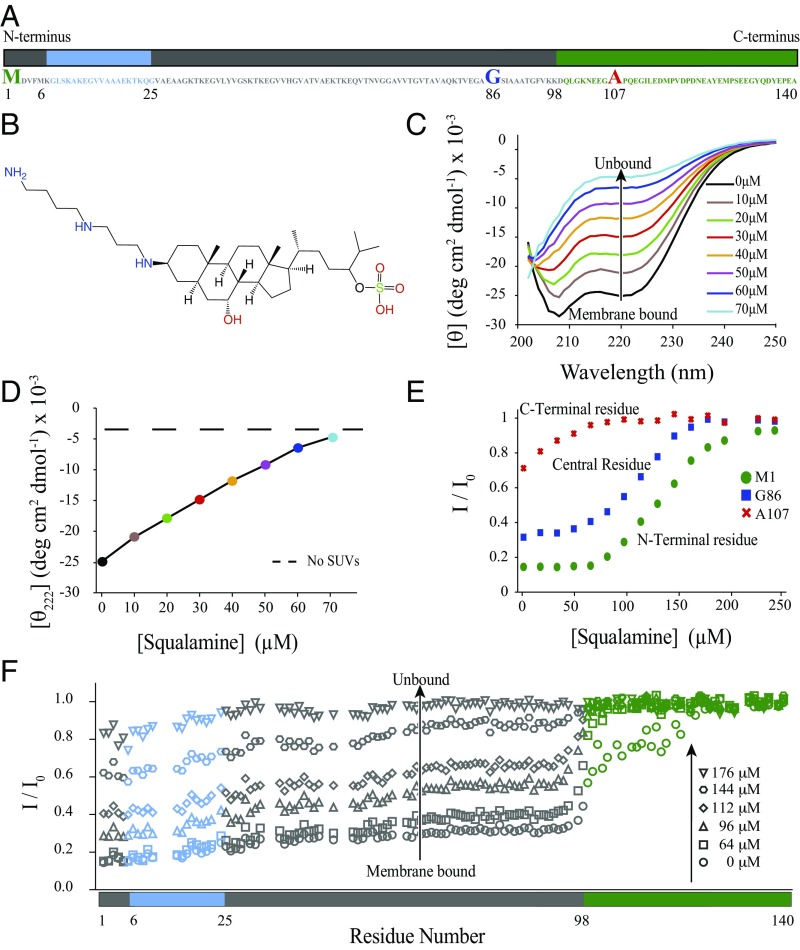
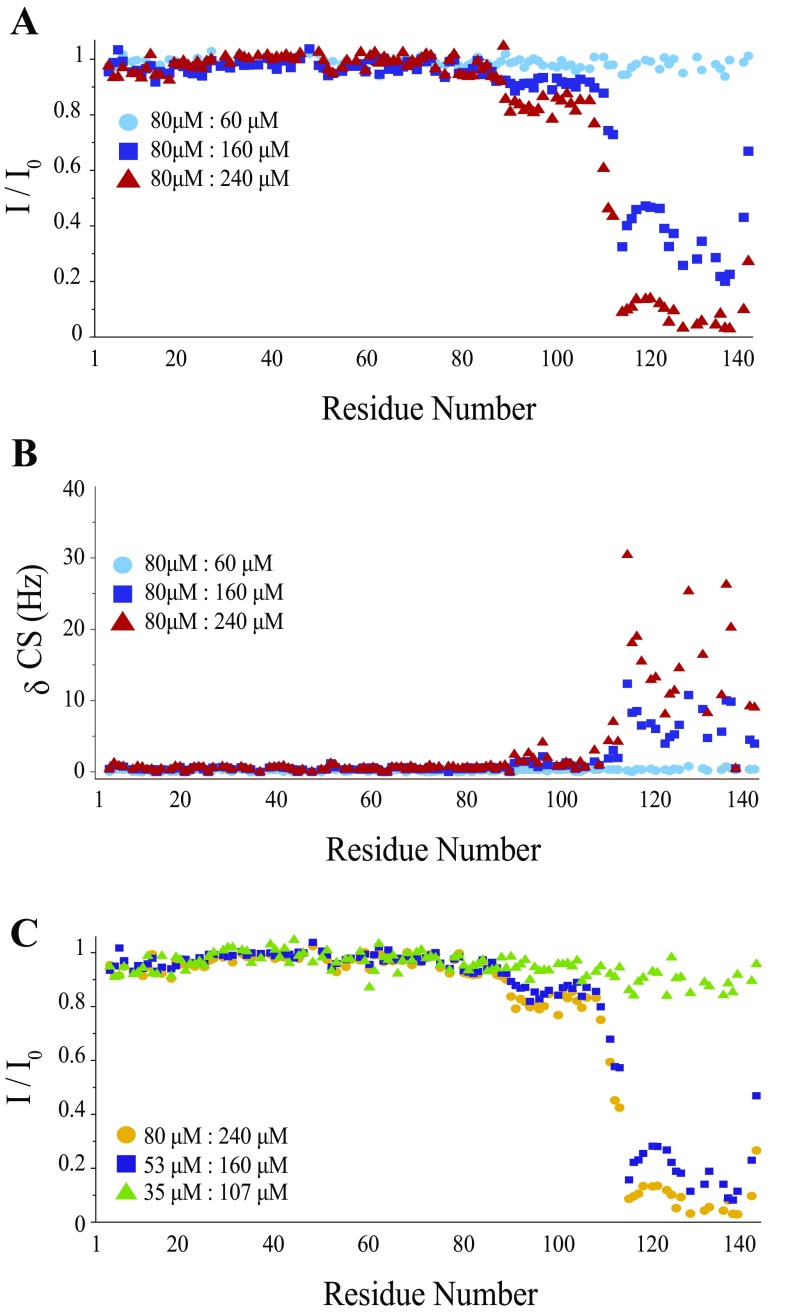
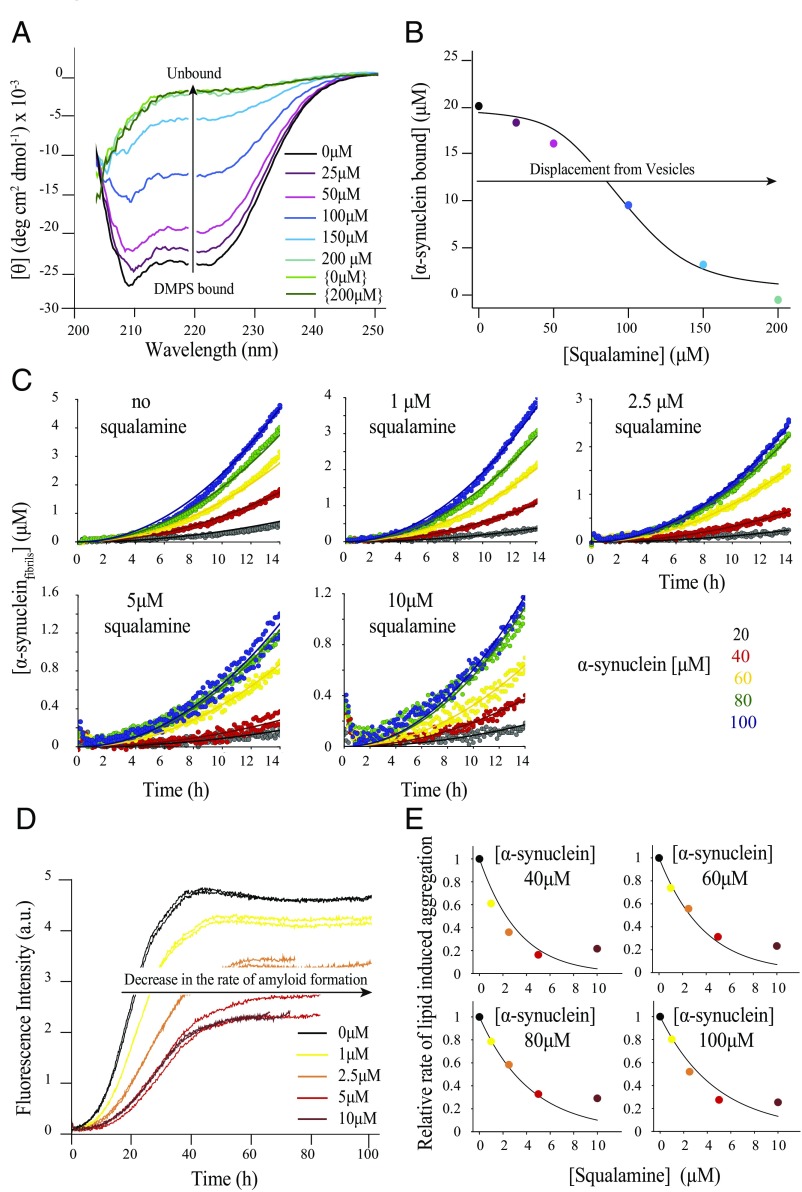
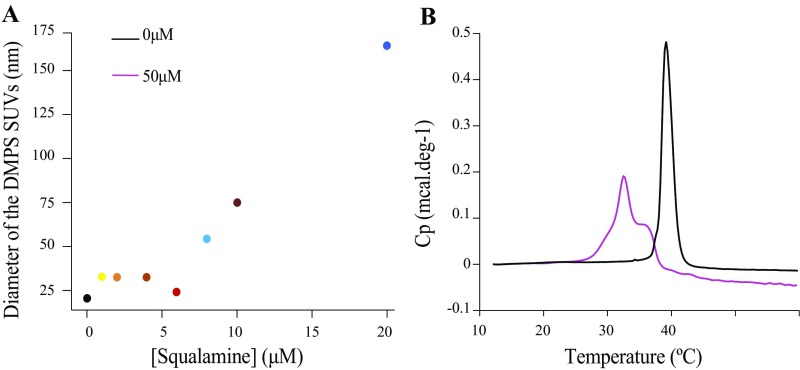
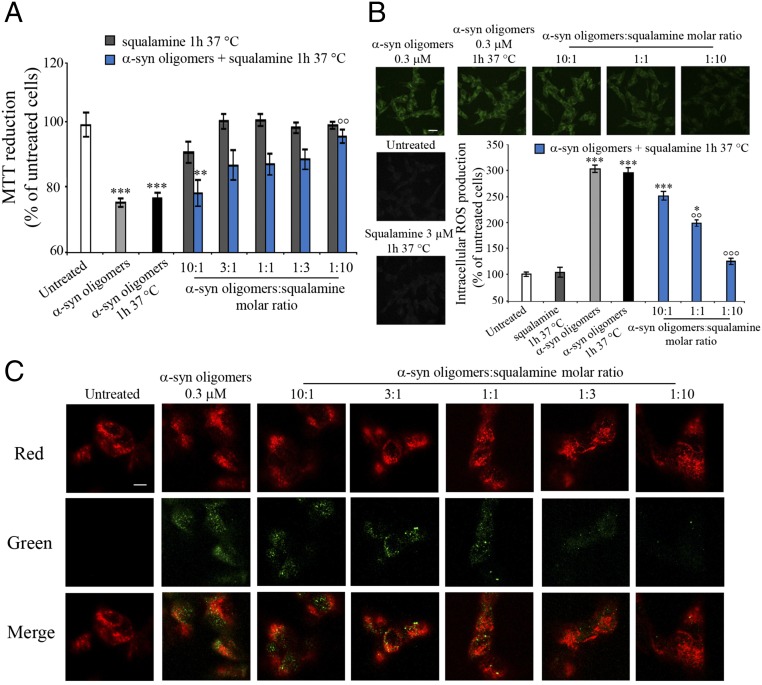
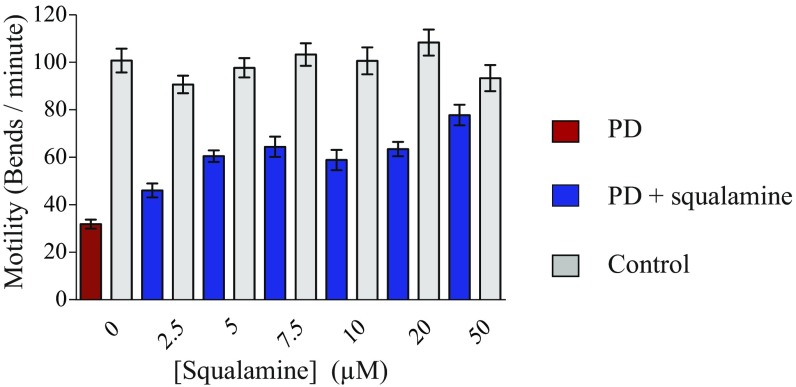
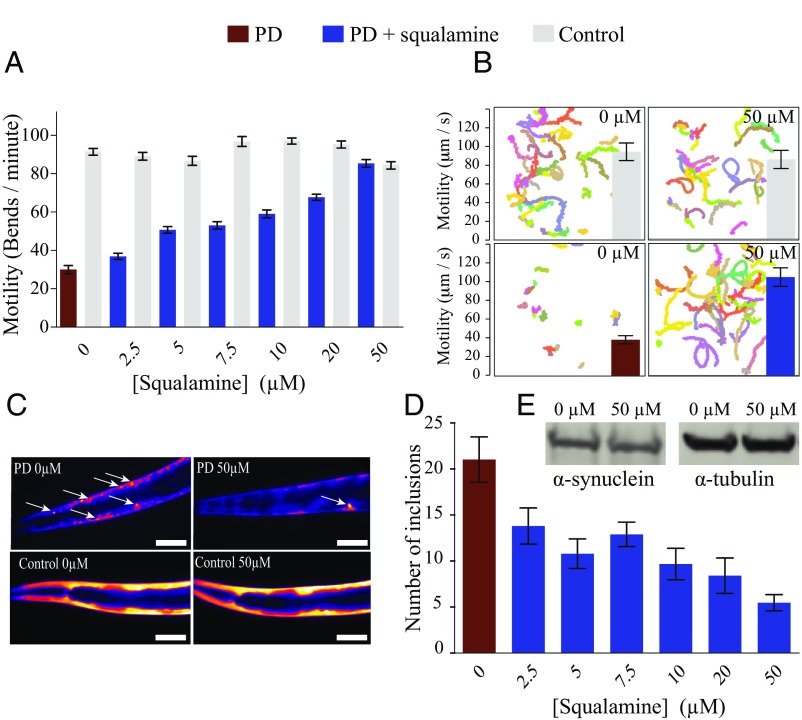
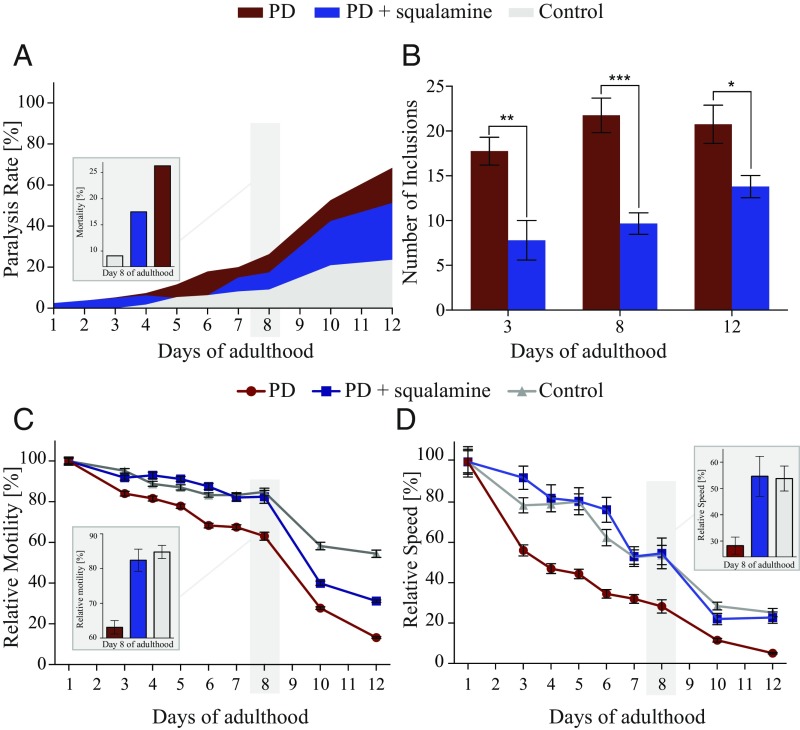
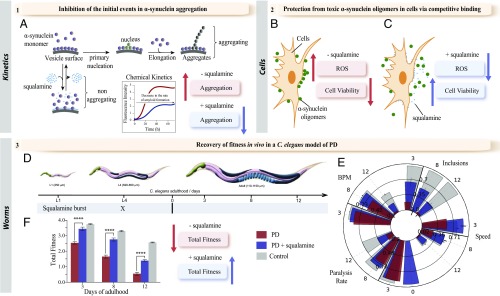
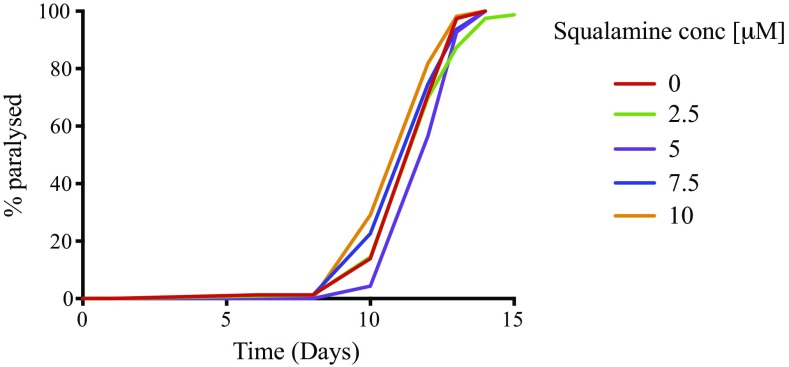
The self-assembly of α-synuclein is closely associated with Parkinson's disease and related syndromes. We show that squalamine, a natural product with known anticancer and antiviral activity, dramatically affects α-synuclein aggregation in vitro and in vivo. We elucidate the mechanism of action of squalamine by investigating its interaction with lipid vesicles, which are known to stimulate nucleation, and find that this compound displaces α-synuclein from the surfaces of such vesicles, thereby blocking the first steps in its aggregation process. We also show that squalamine almost completely suppresses the toxicity of α-synuclein oligomers in human neuroblastoma cells by inhibiting their interactions with lipid membranes. We further examine the effects of squalamine in a Caenorhabditis elegans strain overexpressing α-synuclein, observing a dramatic reduction of α-synuclein aggregation and an almost complete elimination of muscle paralysis. These findings suggest that squalamine could be a means of therapeutic intervention in Parkinson's disease and related conditions.
Keywords: Parkinson’s disease; amyloid formation; drug development; protein aggregation; toxic oligomers.
Conflict of interest statement
M.Z. is the inventor on a patent application that has been filed related to the compound described in this paper. The other authors declare no conflict of interest.
Figures

Comment in
-
Modulating membrane binding of α-synuclein as a therapeutic strategy.Proc Natl Acad Sci U S A. 2017 Feb 7;114(6):1223-1225. doi: 10.1073/pnas.1620159114. Epub 2017 Jan 26. Proc Natl Acad Sci U S A. 2017. PMID: 28126719 Free PMC article. No abstract available.
References
-
- Breydo L, Wu JW, Uversky VN. Α-synuclein misfolding and Parkinson’s disease. Biochim Biophys Acta. 2012;1822(2):261–285. - PubMed
-
- Dettmer U, Selkoe D, Bartels T. New insights into cellular α-synuclein homeostasis in health and disease. Curr Opin Neurobiol. 2016;36:15–22. - PubMed
-
- Dawson TM, Dawson VL. Molecular pathways of neurodegeneration in Parkinson’s disease. Science. 2003;302(5646):819–822. - PubMed
-
- Knowles TPJ, Vendruscolo M, Dobson CM. The amyloid state and its association with protein misfolding diseases. Nat Rev Mol Cell Biol. 2014;15(6):384–396. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
