

Continual renewal and replication of persistent Leishmania major parasites in concomitantly immune hosts
- PMID: 28096392
- PMCID: PMC5293024
- DOI: 10.1073/pnas.1619265114
Continual renewal and replication of persistent Leishmania major parasites in concomitantly immune hosts
Abstract
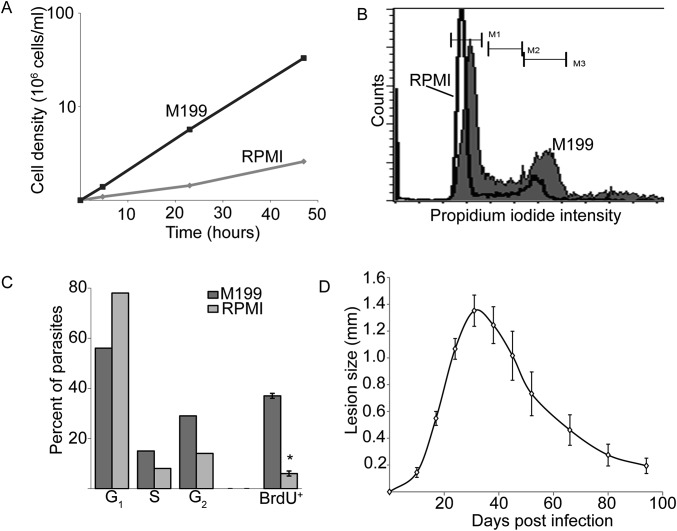
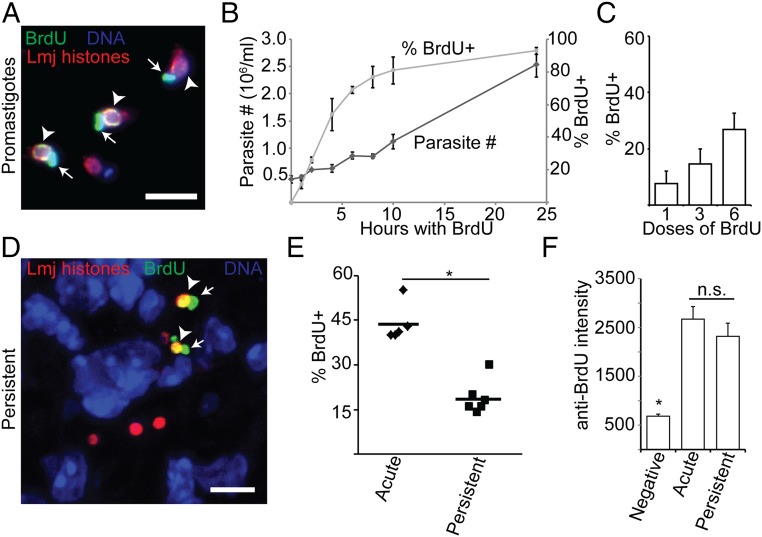
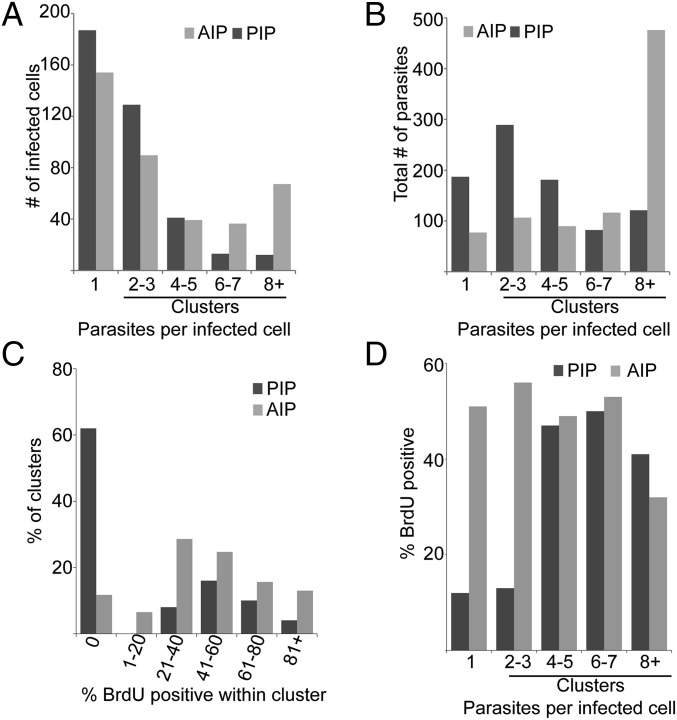
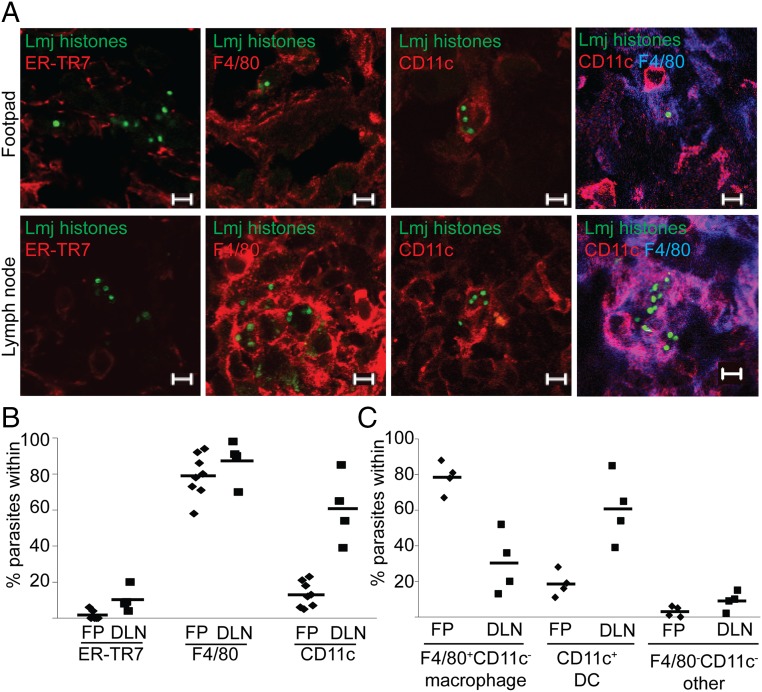
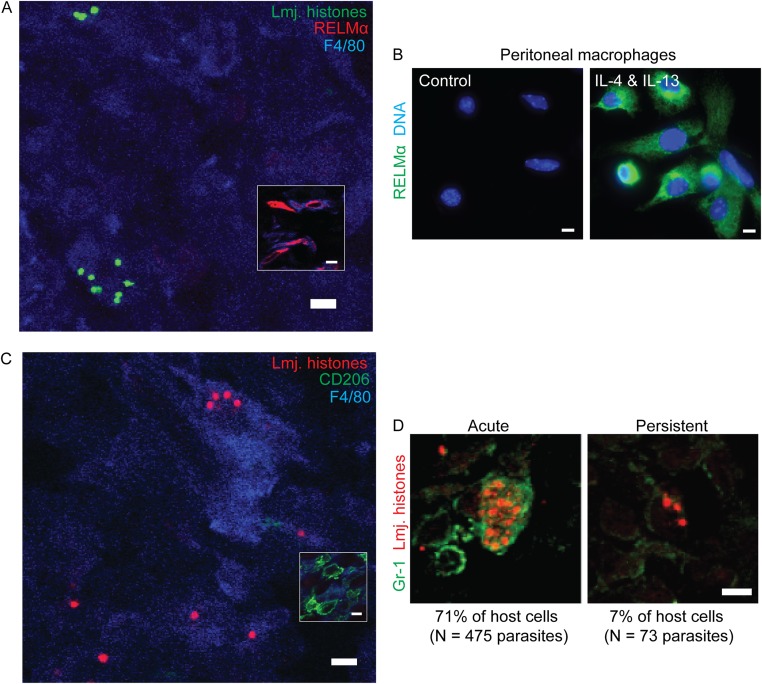
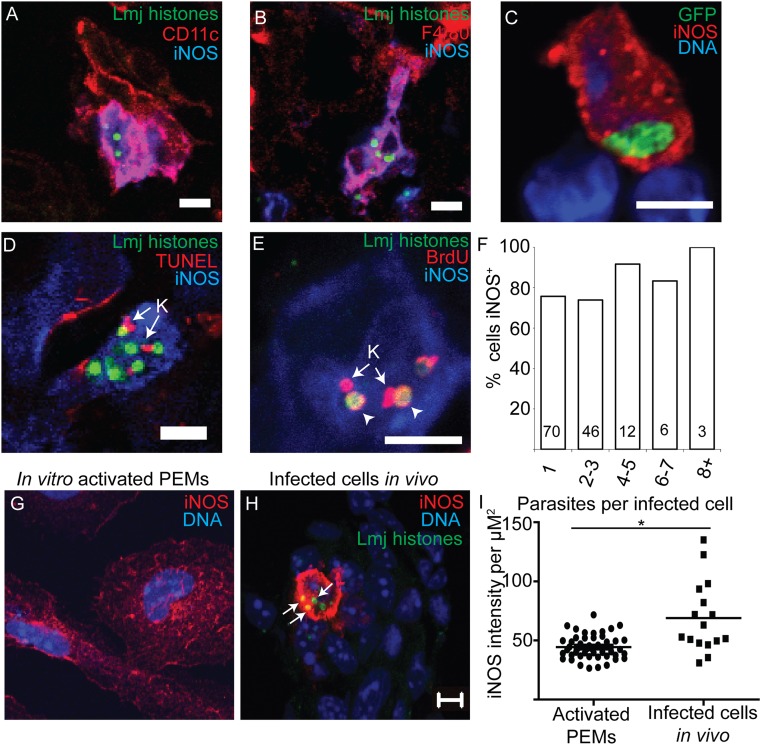
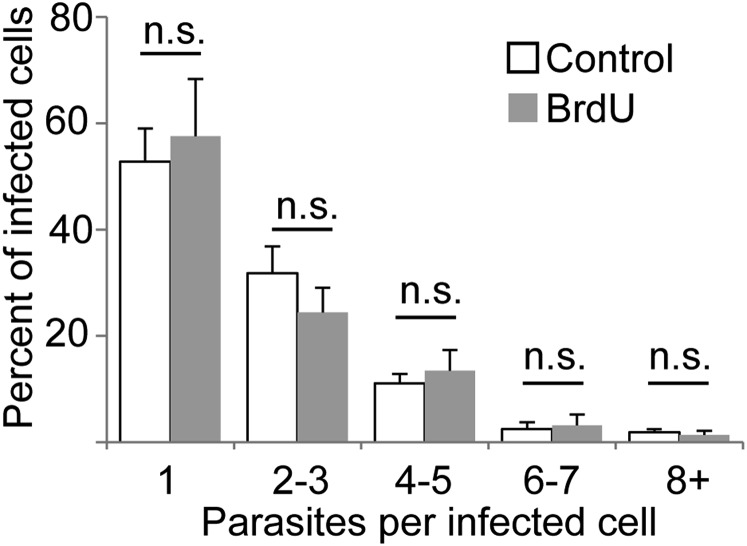
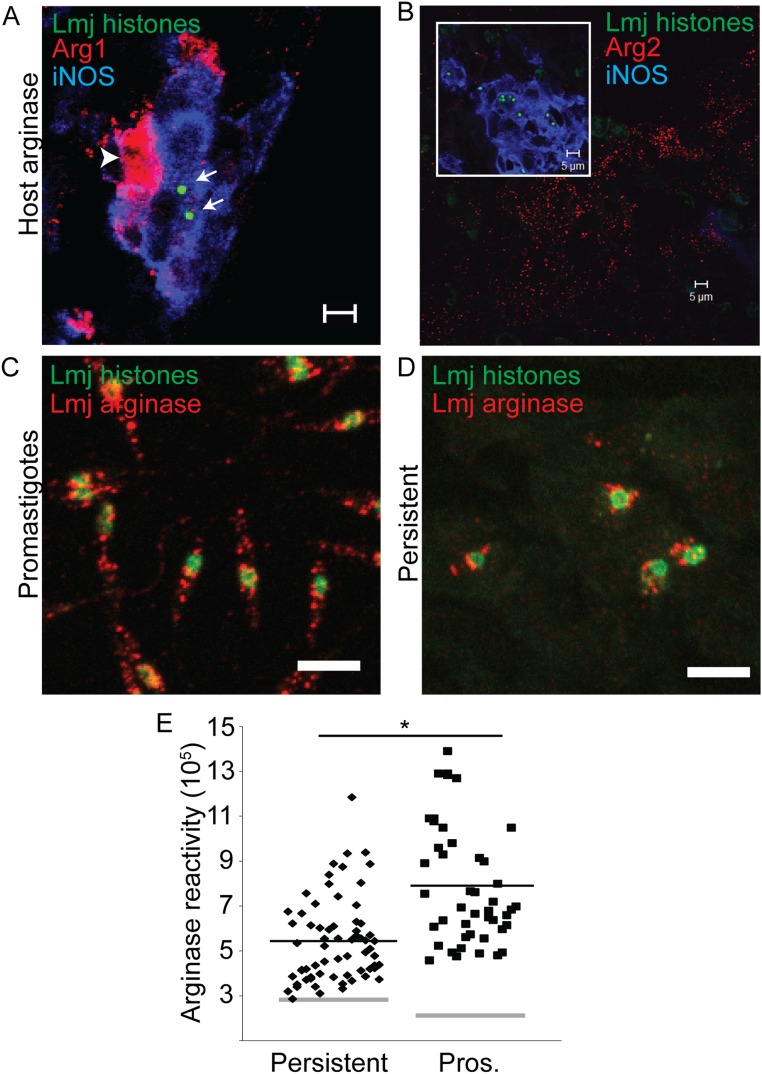
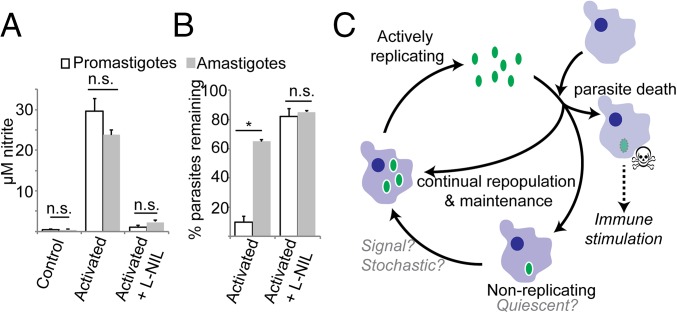
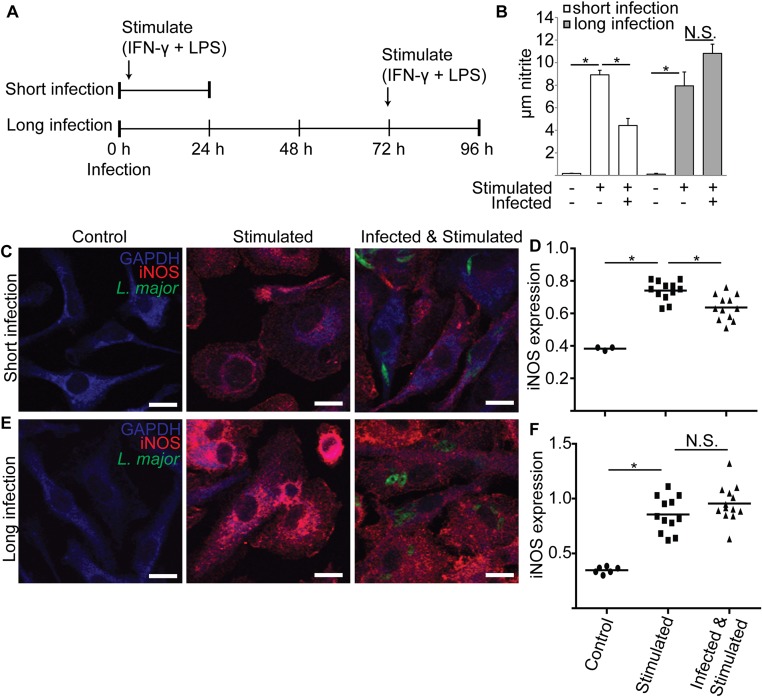
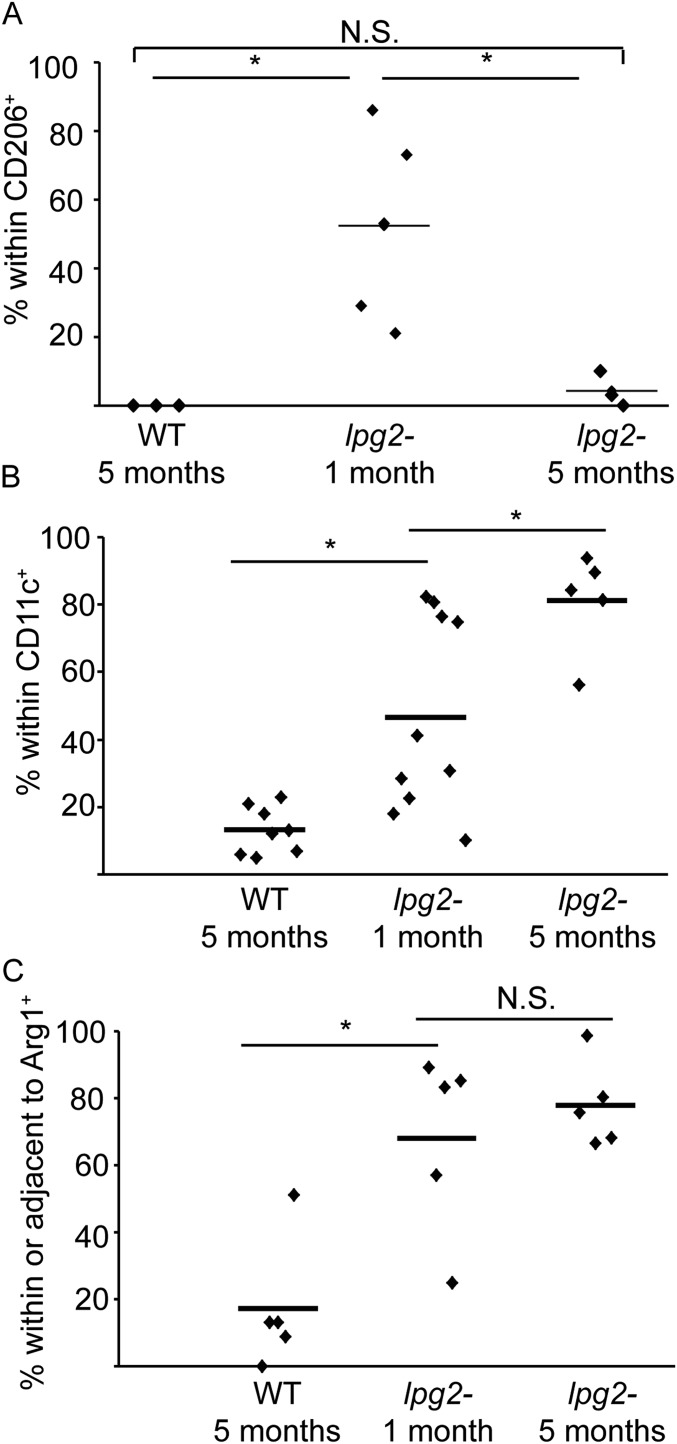
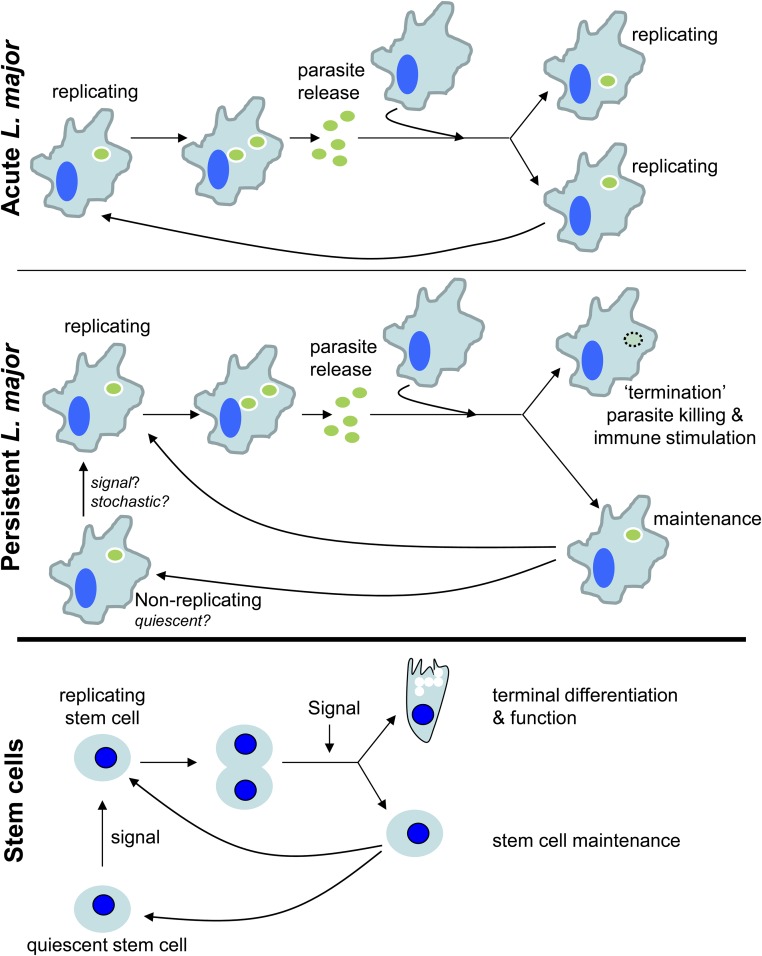
In most natural infections or after recovery, small numbers of Leishmania parasites remain indefinitely in the host. Persistent parasites play a vital role in protective immunity against disease pathology upon reinfection through the process of concomitant immunity, as well as in transmission and reactivation, yet are poorly understood. A key question is whether persistent parasites undergo replication, and we devised several approaches to probe the small numbers in persistent infections. We find two populations of persistent Leishmania major: one rapidly replicating, similar to parasites in acute infections, and another showing little evidence of replication. Persistent Leishmania were not found in "safe" immunoprivileged cell types, instead residing in macrophages and DCs, ∼60% of which expressed inducible nitric oxide synthase (iNOS). Remarkably, parasites within iNOS+ cells showed normal morphology and genome integrity and labeled comparably with BrdU to parasites within iNOS- cells, suggesting that these parasites may be unexpectedly resistant to NO. Nonetheless, because persistent parasite numbers remain roughly constant over time, their replication implies that ongoing destruction likewise occurs. Similar results were obtained with the attenuated lpg2- mutant, a convenient model that rapidly enters a persistent state without inducing pathology due to loss of the Golgi GDP mannose transporter. These data shed light on Leishmania persistence and concomitant immunity, suggesting a model wherein a parasite reservoir repopulates itself indefinitely, whereas some progeny are terminated in antigen-presenting cells, thereby stimulating immunity. This model may be relevant to understanding immunity to other persistent pathogen infections.
Keywords: latency; quiescence; stem cell-like; trypanosomatid protozoan parasite; vaccination.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Monack DM, Mueller A, Falkow S. Persistent bacterial infections: The interface of the pathogen and the host immune system. Nat Rev Microbiol. 2004;2(9):747–765. - PubMed
-
- Virgin HW, Wherry EJ, Ahmed R. Redefining chronic viral infection. Cell. 2009;138(1):30–50. - PubMed
-
- Roizman B, Whitley RJ. An inquiry into the molecular basis of HSV latency and reactivation. Annu Rev Microbiol. 2013;67:355–374. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
