

Trogocytosis of peptide-MHC class II complexes from dendritic cells confers antigen-presenting ability on basophils
- PMID: 28096423
- PMCID: PMC5293035
- DOI: 10.1073/pnas.1615973114
Trogocytosis of peptide-MHC class II complexes from dendritic cells confers antigen-presenting ability on basophils
Abstract
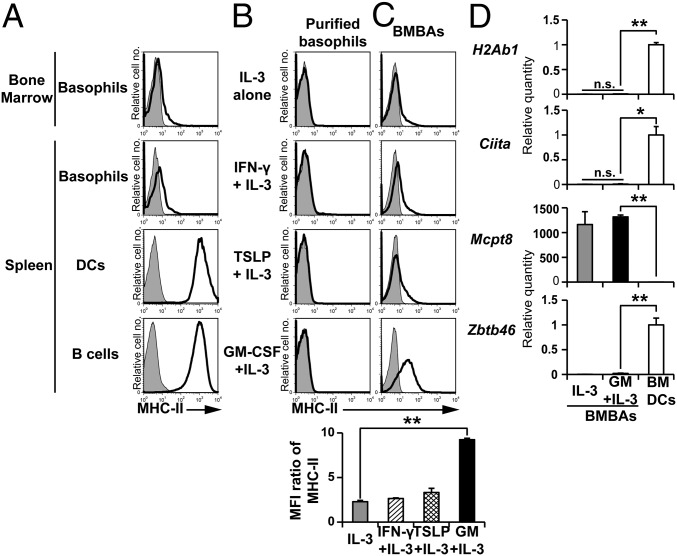
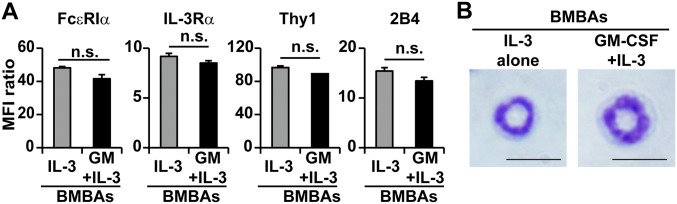
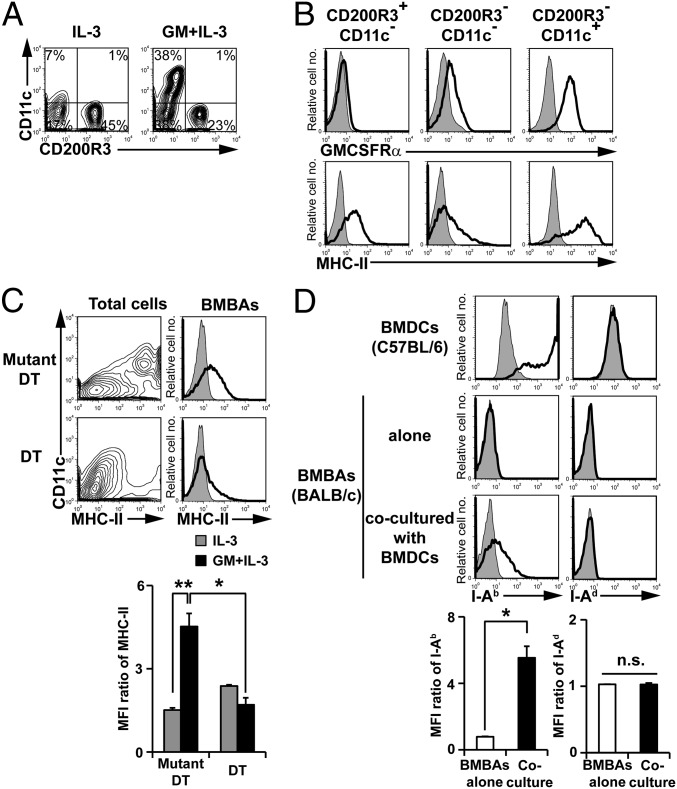
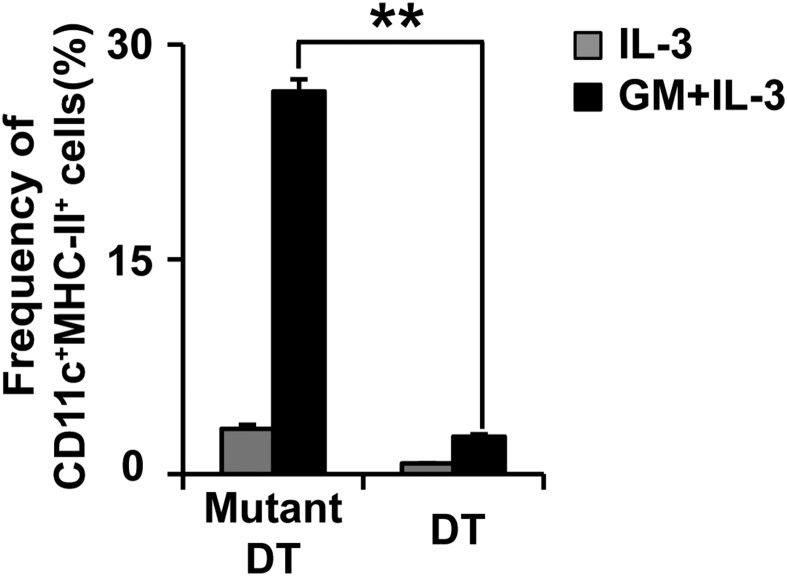
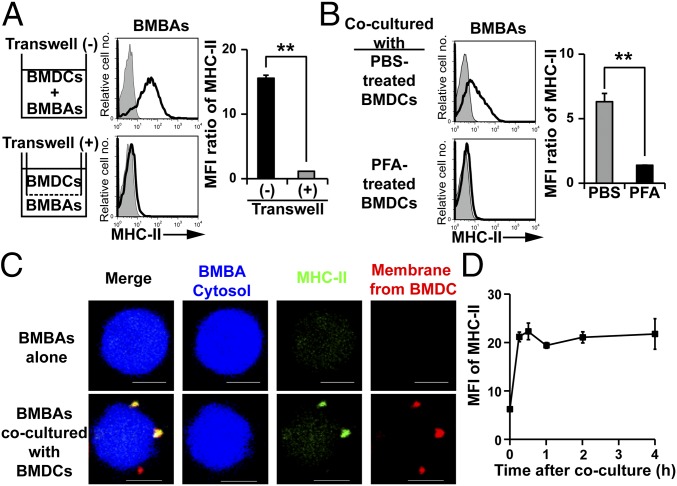
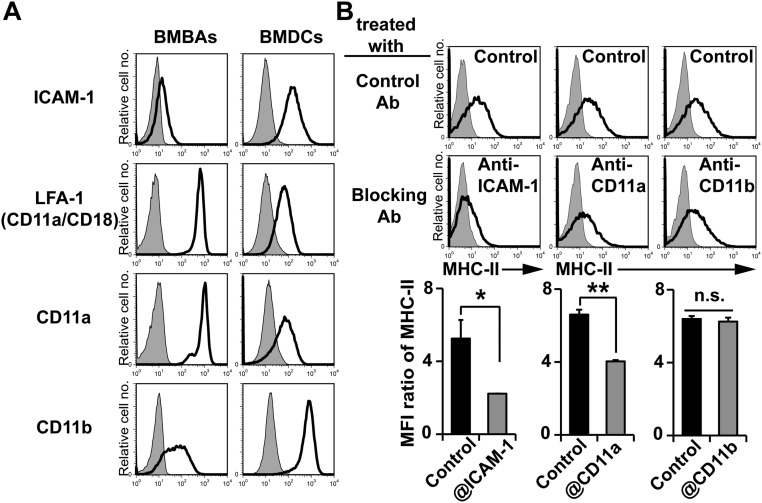
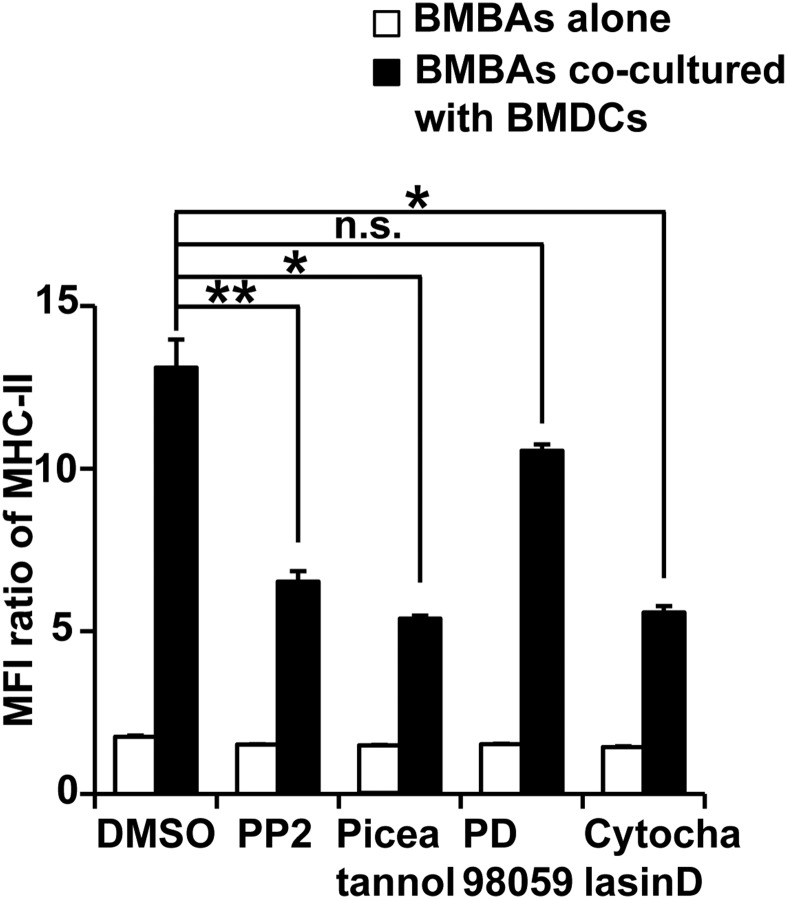
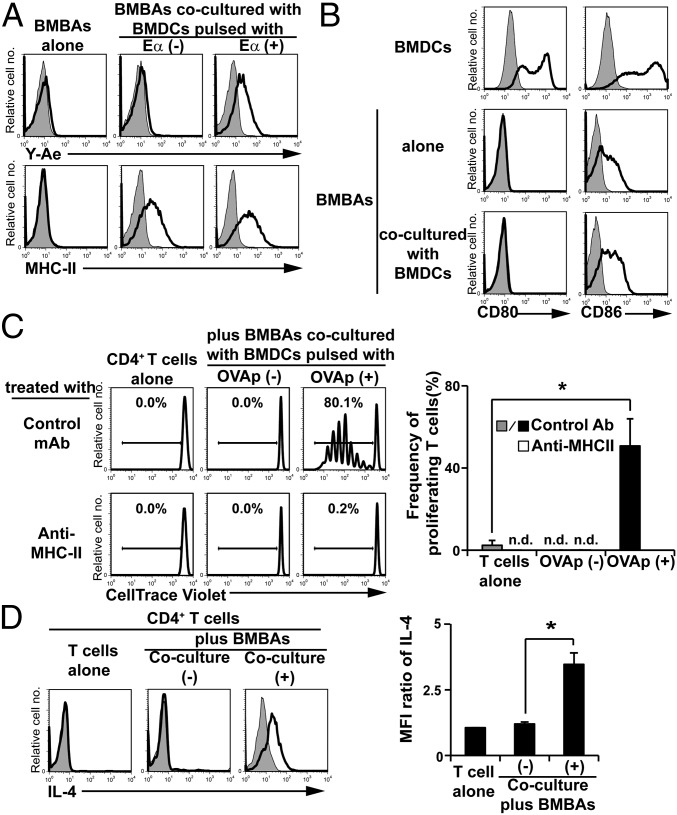
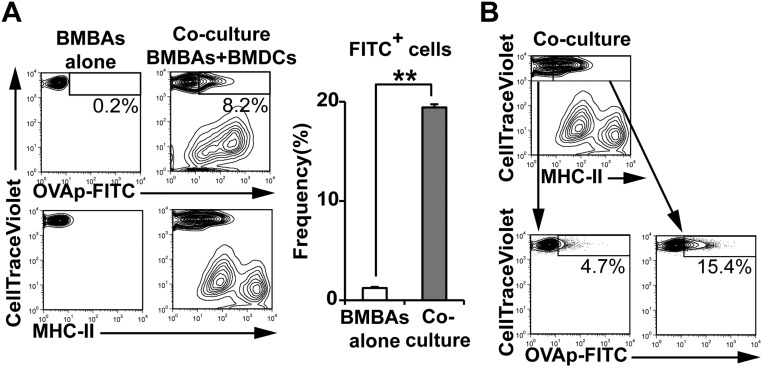
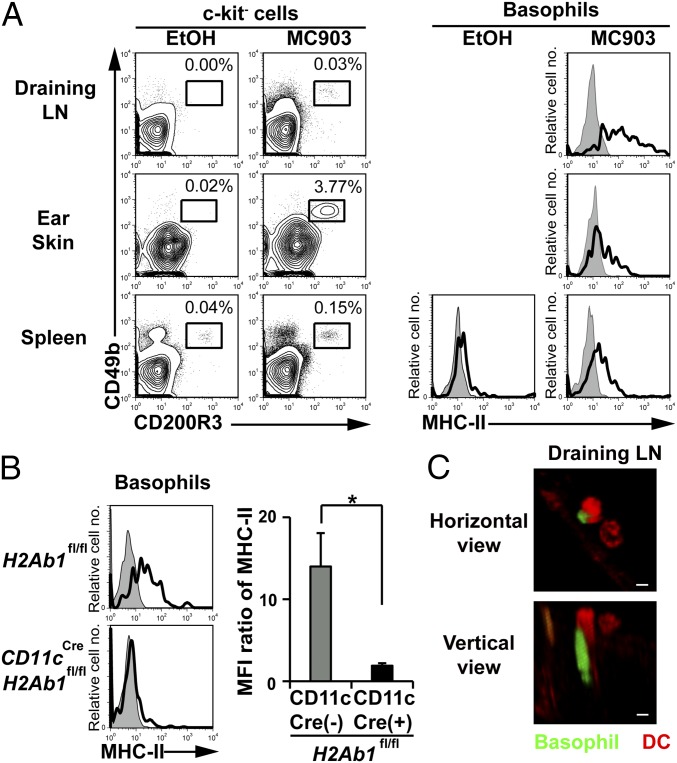
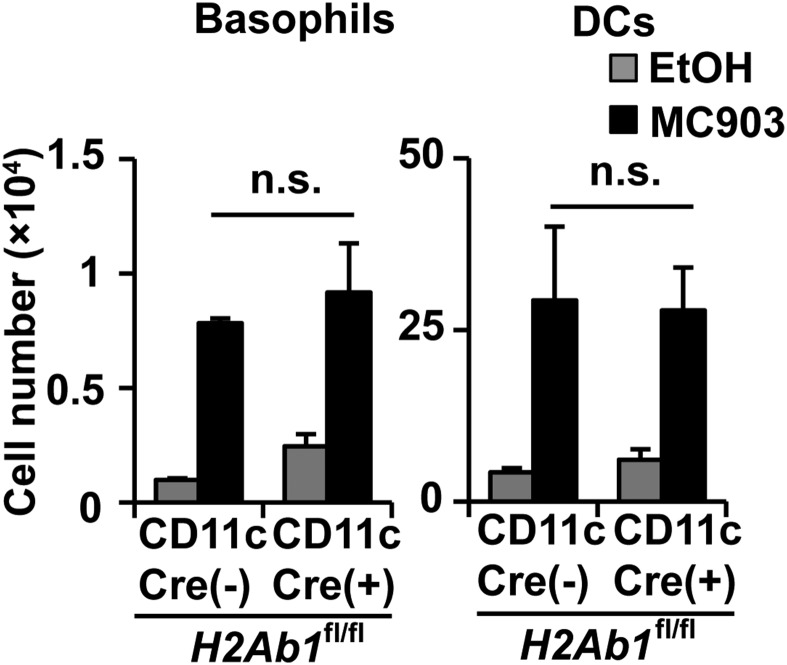
Th2 immunity plays important roles in both protective and allergic responses. Nevertheless, the nature of antigen-presenting cells responsible for Th2 cell differentiation remains ill-defined compared with the nature of the cells responsible for Th1 and Th17 cell differentiation. Basophils have attracted attention as a producer of Th2-inducing cytokine IL-4, whereas their MHC class II (MHC-II) expression and function as antigen-presenting cells are matters of considerable controversy. Here we revisited the MHC-II expression on basophils and explored its functional relevance in Th2 cell differentiation. Basophils generated in vitro from bone marrow cells in culture with IL-3 plus GM-CSF displayed MHC-II on the cell surface, whereas those generated in culture with IL-3 alone did not. Of note, these MHC-II-expressing basophils showed little or no transcription of the corresponding MHC-II gene. The GM-CSF addition to culture expanded dendritic cells (DCs) other than basophils. Coculture of basophils and DCs revealed that basophils acquired peptide-MHC-II complexes from DCs via cell contact-dependent trogocytosis. The acquired complexes, together with CD86, enabled basophils to stimulate peptide-specific T cells, leading to their proliferation and IL-4 production, indicating that basophils can function as antigen-presenting cells for Th2 cell differentiation. Transfer of MHC-II from DCs to basophils was also detected in draining lymph nodes of mice with atopic dermatitis-like skin inflammation. Thus, the present study defined the mechanism by which basophils display MHC-II on the cell surface and appears to reconcile some discrepancies observed in previous studies.
Keywords: MHC class II; basophil; dendritic cell; trogocytosis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Galli SJ. Mast cells and basophils. Curr Opin Hematol. 2000;7(1):32–39. - PubMed
-
- Sokol CL, Medzhitov R. Emerging functions of basophils in protective and allergic immune responses. Mucosal Immunol. 2010;3(2):129–137. - PubMed
-
- Karasuyama H, Mukai K, Obata K, Tsujimura Y, Wada T. Nonredundant roles of basophils in immunity. Annu Rev Immunol. 2011;29:45–69. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
