

Regulation of aldosterone secretion by mineralocorticoid receptor-mediated signaling
- PMID: 28096435
- PMCID: PMC5464000
- DOI: 10.1530/JOE-16-0452
Regulation of aldosterone secretion by mineralocorticoid receptor-mediated signaling
Abstract
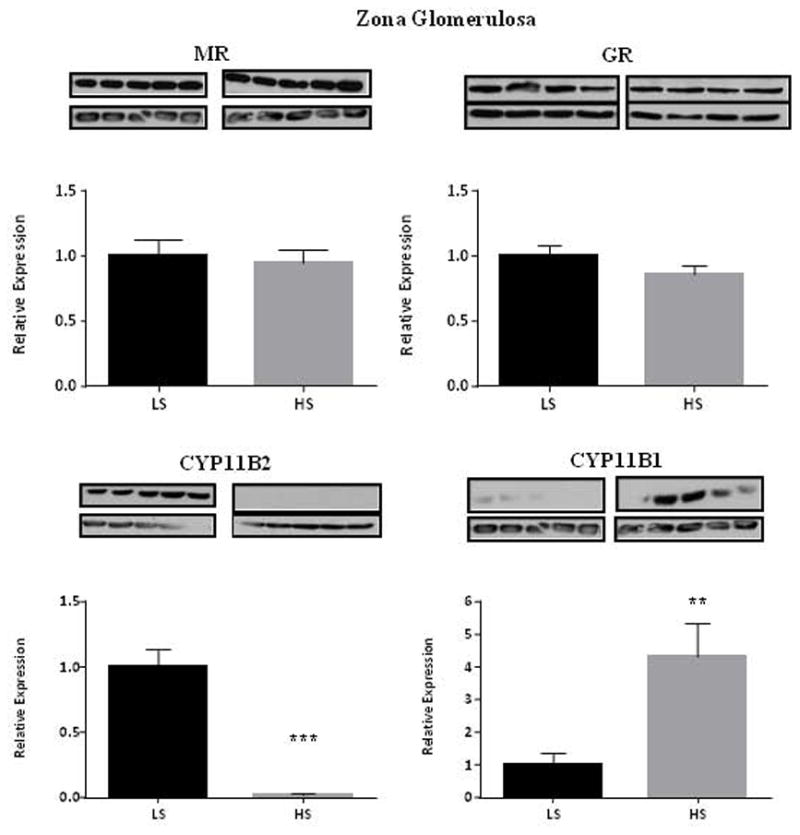
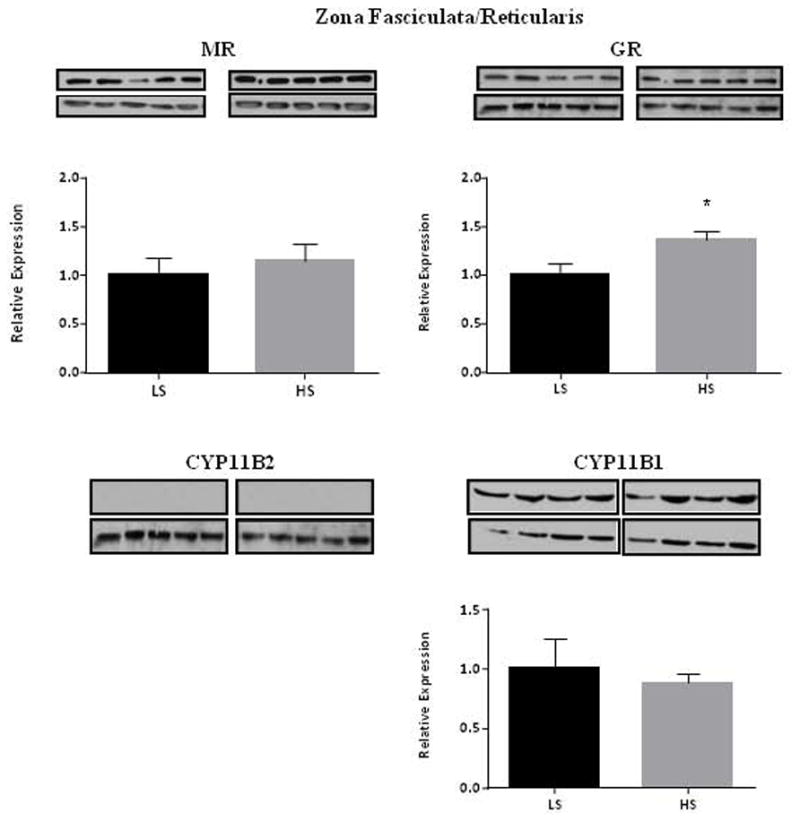
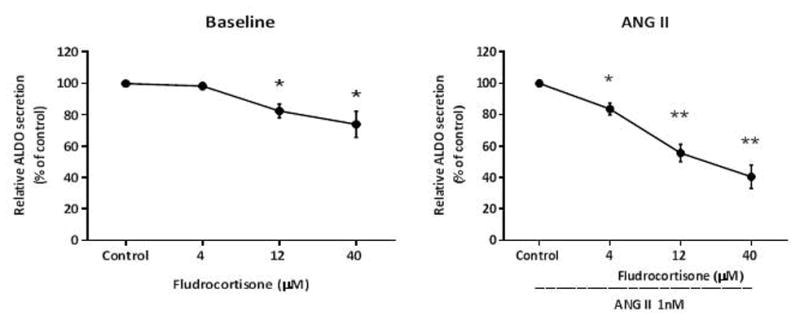
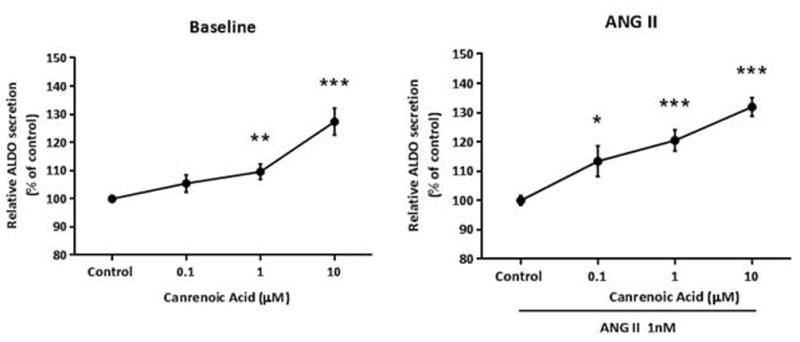
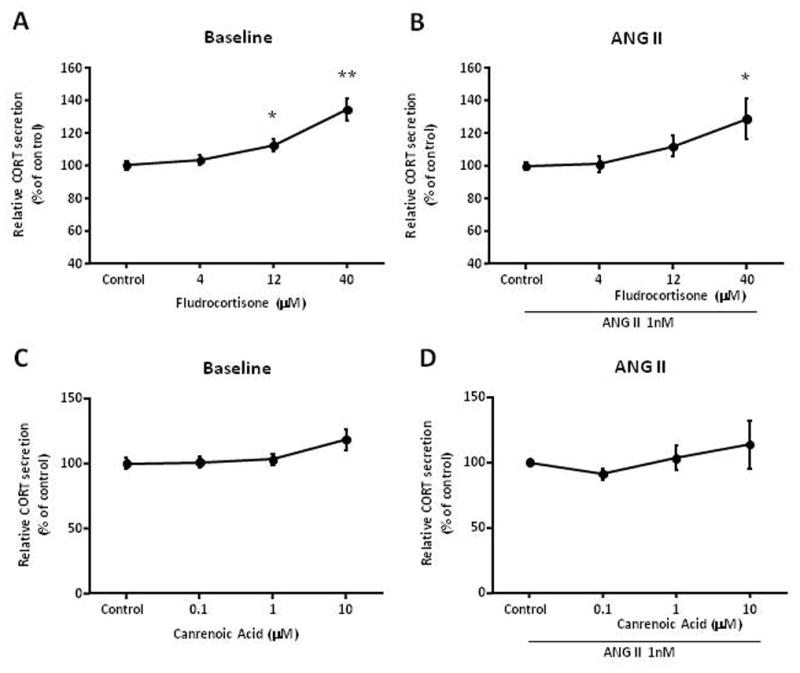
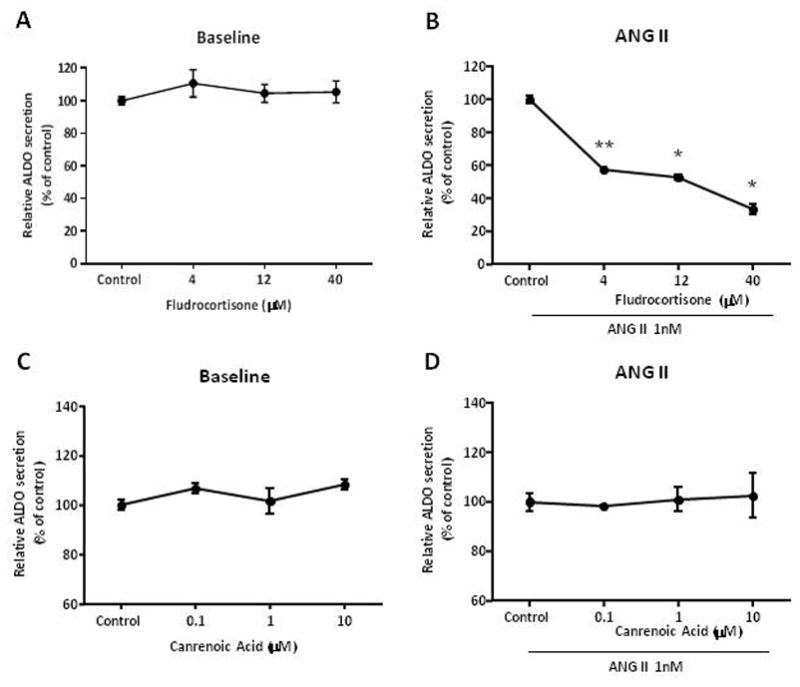
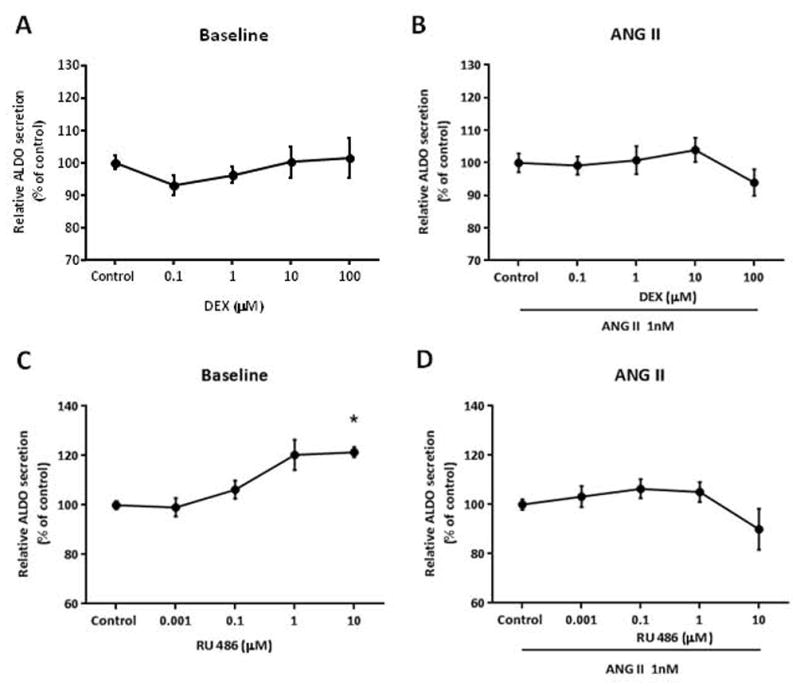
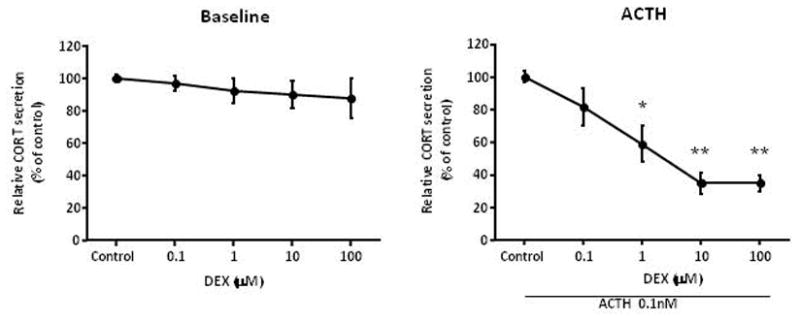
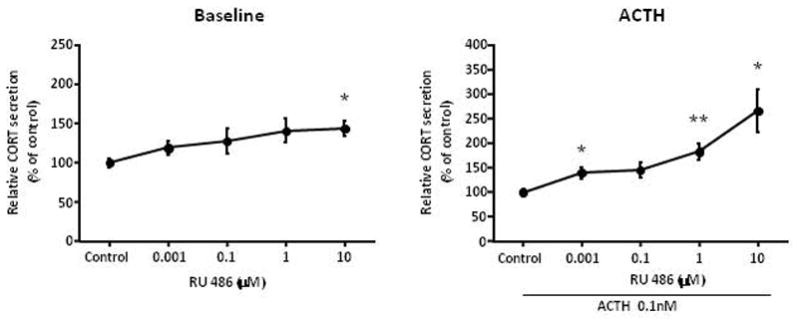
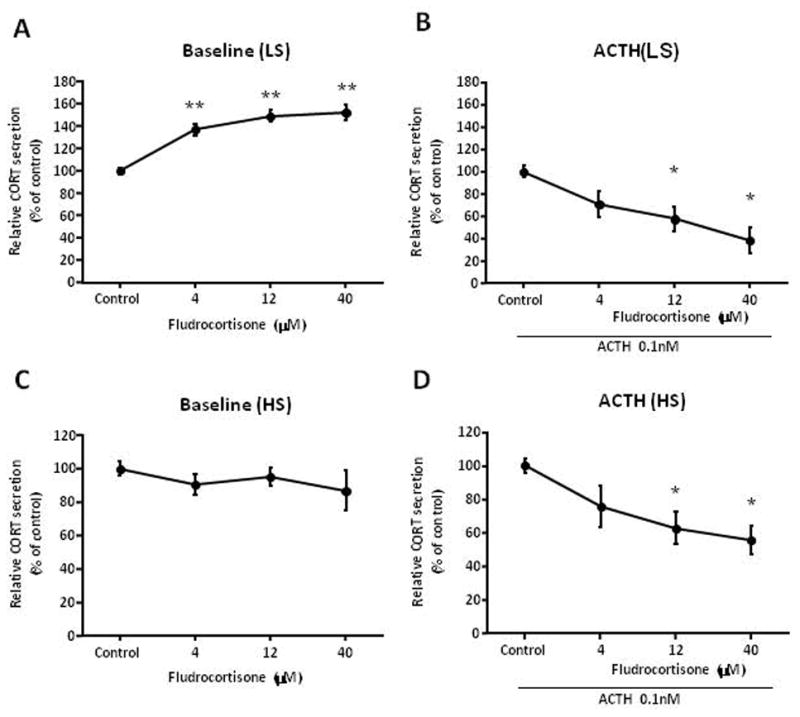
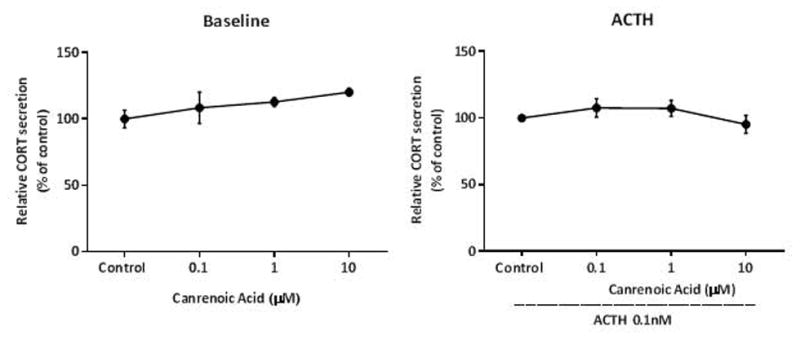
We posit the existence of a paracrine/autocrine negative feedback loop, mediated by the mineralocorticoid receptor (MR), regulating aldosterone secretion. To assess this hypothesis, we asked whether altering MR activity in zona glomerulosa (ZG) cells affects aldosterone production. To this end, we studied ex vivo ZG cells isolated from male Wistar rats fed chow containing either high (1.6% Na+ (HS)) or low (0.03% Na+ (LS)) amount of sodium. Western blot analyses demonstrated that MR was present in both the ZG and zona fasciculata/zona reticularis (ZF/ZR/ZR). In ZG cells isolated from rats on LS chow, MR activation by fludrocortisone produced a 20% and 60% reduction in aldosterone secretion basally and in response to angiotensin II (ANGII) stimulation, respectively. Corticosterone secretion was increased in these cells suggesting that aldosterone synthase activity was being reduced by fludrocortisone. In contrast, canrenoic acid, an MR antagonist, enhanced aldosterone production by up to 30% both basally and in response to ANGII. Similar responses were observed in ZG cells from rats fed HS. Modulating glucocorticoid receptor (GR) activity did not alter aldosterone production by ZG cells; however, altering GR activity did modify corticosterone production from ZF/ZR/ZR cells both basally and in response to adrenocorticotropic hormone (ACTH). Additionally, activating the MR in ZF/ZR/ZR cells strikingly reduced corticosterone secretion. In summary, these data support the hypothesis that negative ultra-short feedback loops regulate adrenal steroidogenesis. In the ZG, aldosterone secretion is regulated by the MR, but not the GR, an effect that appears to be secondary to a change in aldosterone synthase activity.
Keywords: adrenal cortex; aldosterone; corticosterone; zona fasciculata; zona glomerulosa.
© 2017 Society for Endocrinology.
Figures
References
-
- Agarwal MK, Coupry F, Philippe M. Physiological activity and receptor binding of 9 alpha fluorohydrocortisone. Biochem Biophys Res Commun. 1977;78(2):747–753. - PubMed
-
- Aguilera G, Catt KJ. Loci of action of regulators of aldosterone biosynthesis in isolated glomerulosa cells. Endocrinology. 1979;104(4):1046–1052. - PubMed
-
- Arvat E, Maccagno B, Giordano R, Pellegrino M, Broglio F, Gianotti L, Maccario M, Camanni F, Ghigo E. Mineralocorticoid receptor blockade by canrenoate increases both spontaneous and stimulated adrenal function in humans. J Clin Endocrinol Metab. 2001;86(7):3176–3181. - PubMed
-
- Asser L, Hescot S, Viengchareun S, Delemer B, Trabado S, Lombes M. Autocrine positive regulatory feedback of glucocorticoid secretion: glucocorticoid receptor directly impacts H295R human adrenocortical cell function. Mol Cell Endocrinol. 2014;395(1–2):1–9. - PubMed
-
- Baird A, Kan KW, Solomon S. Androstenedione-mediated inhibition of 11 beta-hydroxylation in monolayer cell cultures of fetal calf adrenals. J Steroid Biochem. 1983;18(5):581–584. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
