

Tryptophan metabolite activation of the aryl hydrocarbon receptor regulates IL-10 receptor expression on intestinal epithelia
- PMID: 28098246
- PMCID: PMC5515702
- DOI: 10.1038/mi.2016.133
Tryptophan metabolite activation of the aryl hydrocarbon receptor regulates IL-10 receptor expression on intestinal epithelia
Abstract
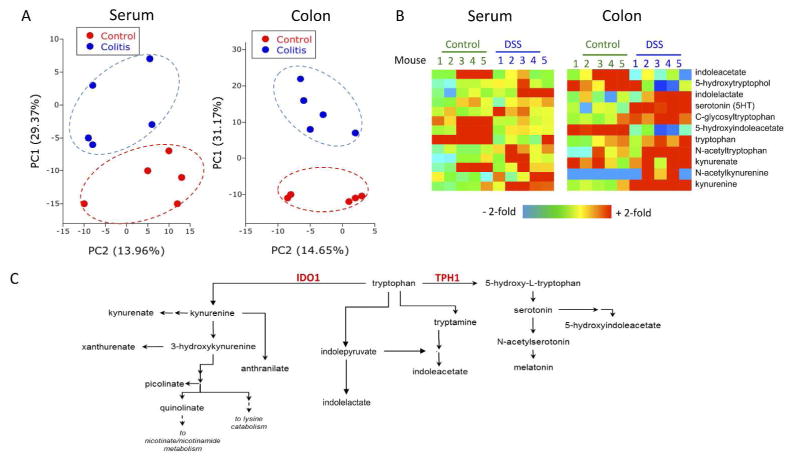
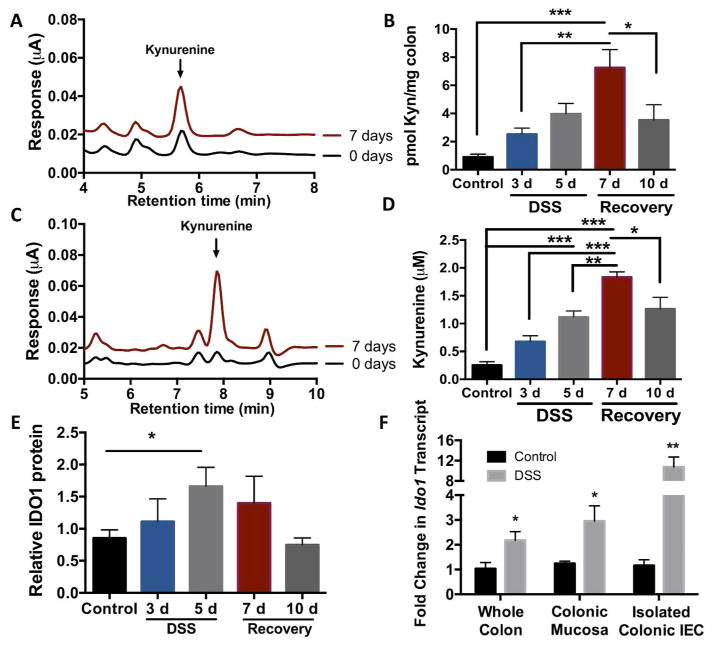
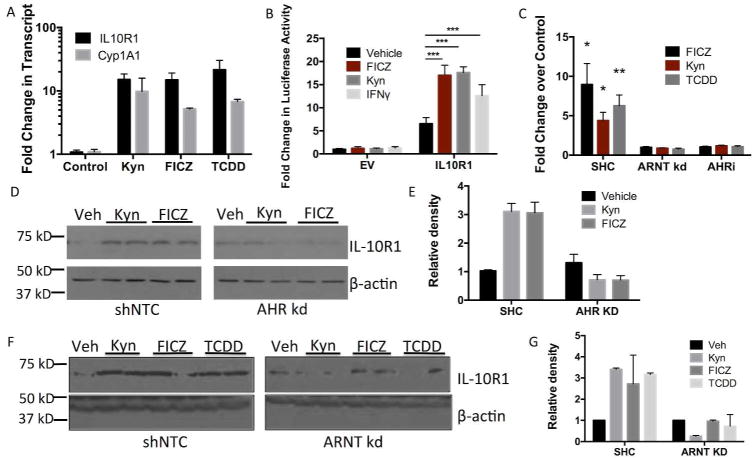
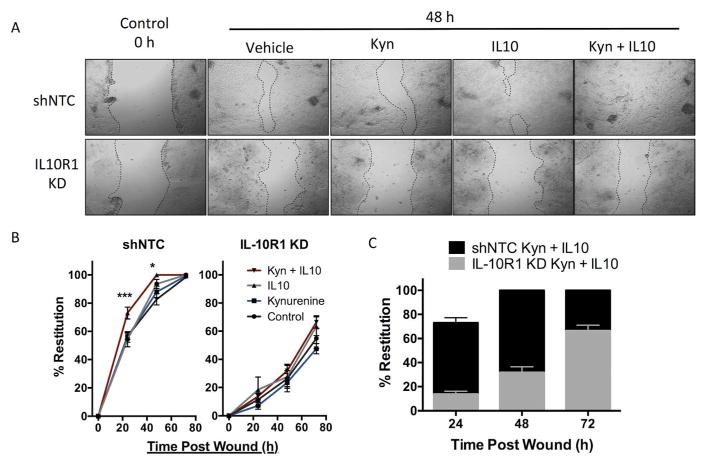
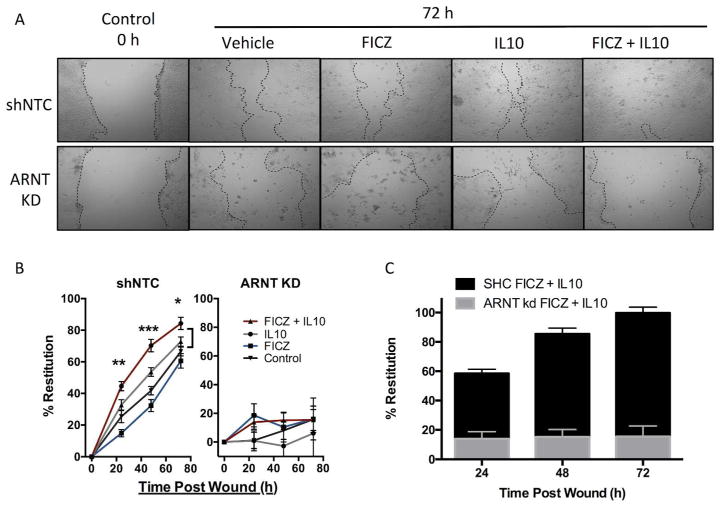
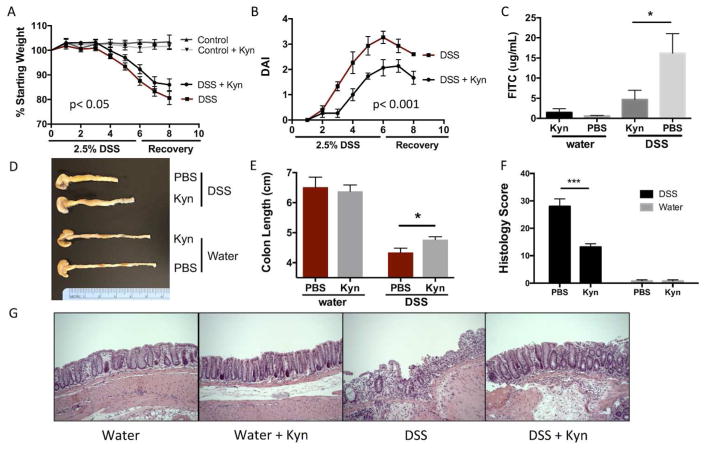
IL-10 is a potent anti-inflammatory cytokine that inhibits the production of proinflammatory mediators. Signaling by IL-10 occurs through the IL-10 receptor (IL-10R), which is expressed in numerous cell types, including intestinal epithelial cells (IECs), where it is associated with development and maintenance of barrier function. Guided by an unbiased metabolomics screen, we identified tryptophan (Trp) metabolism as a major modifying pathway in interferon-γ (IFNγ)-dominant murine colitis. In parallel, we demonstrated that IFNγ induction of indoleamine 2,3-dioxygenase 1, an enzyme that catalyzes the conversion of Trp to kynurenine (Kyn), induces IL-10R1 expression. Based on these findings, we hypothesized that IL-10R1 expression on IEC is regulated by Trp metabolites. Analysis of the promoter region of IL-10R1 revealed a functional aryl hydrocarbon response element, which is induced by Kyn in luciferase-based IL-10R1 promoter assays. Additionally, this analysis confirmed that IL-10R1 protein levels were increased in response to Kyn in IEC in vitro. Studies using in vitro wounding assays revealed that Kyn accelerates IL-10-dependent wound closure. Finally, reduction of murine dextran sodium sulfate colitis through Kyn administration correlates with colonic IL-10R1 expression. Taken together, these results provide evidence on the importance of IL-10 signaling in intestinal epithelia and implicate AHR in the regulation of IL-10R1 expression in the colon.
Conflict of interest statement
Figures
References
-
- Candela M, Perna F, Carnevali P, Vitali B, Ciati R, Gionchetti P, et al. Interaction of probiotic Lactobacillus and Bifidobacterium strains with human intestinal epithelial cells: adhesion properties, competition against enteropathogens and modulation of IL-8 production. International journal of food microbiology. 2008;125(3):286–292. - PubMed
-
- Fukuda S, Toh H, Hase K, Oshima K, Nakanishi Y, Yoshimura K, et al. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature. 2011;469(7331):543–547. - PubMed
-
- Sonnenburg JL, Xu J, Leip DD, Chen CH, Westover BP, Weatherford J, et al. Glycan foraging in vivo by an intestine-adapted bacterial symbiont. Science (New York, NY) 2005;307(5717):1955–1959. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
