

Axon Termination, Pruning, and Synaptogenesis in the Giant Fiber System of Drosophila melanogaster Is Promoted by Highwire
- PMID: 28100586
- PMCID: PMC5340335
- DOI: 10.1534/genetics.116.197343
Axon Termination, Pruning, and Synaptogenesis in the Giant Fiber System of Drosophila melanogaster Is Promoted by Highwire
Abstract
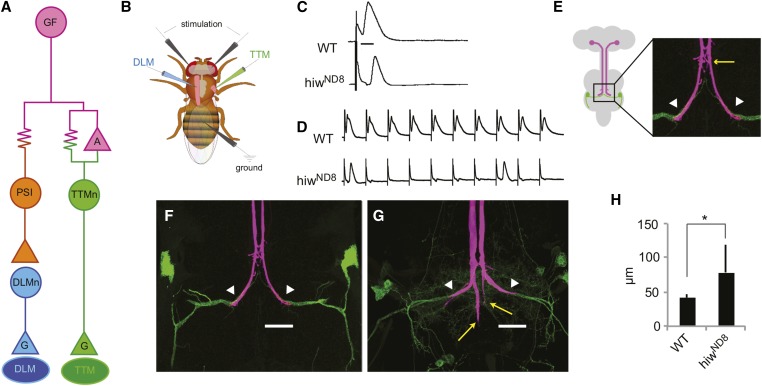
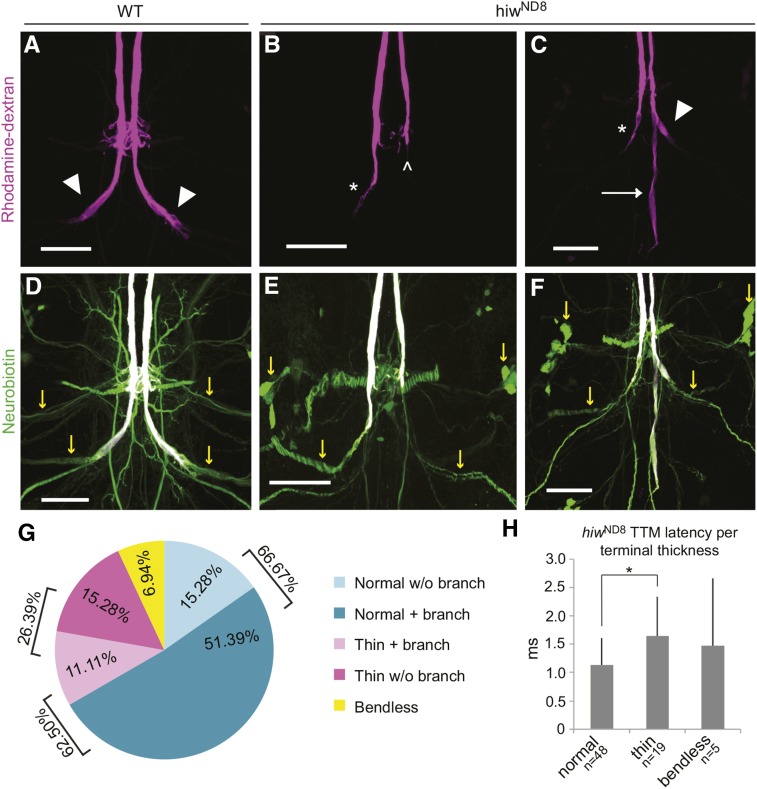
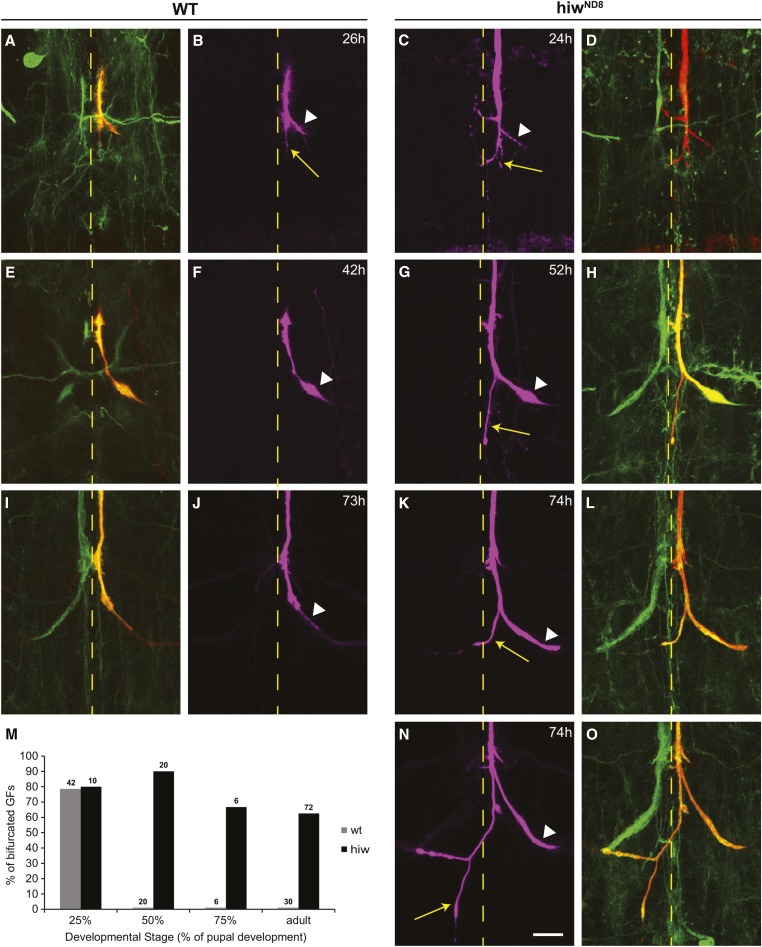
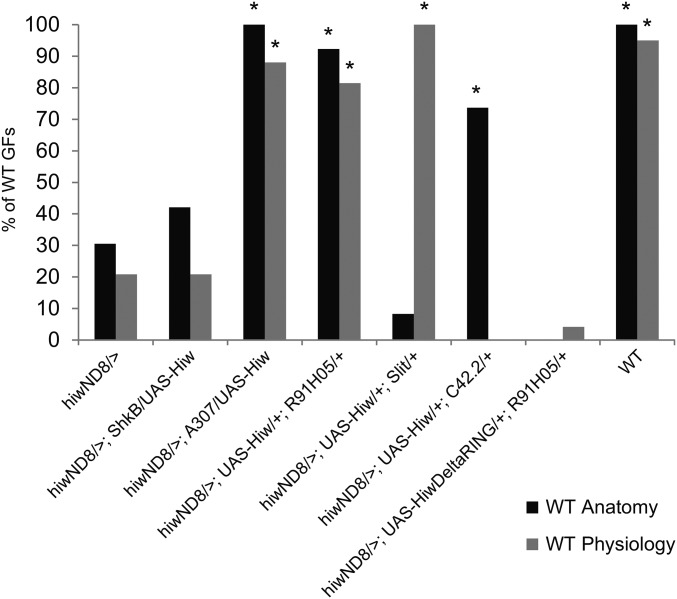
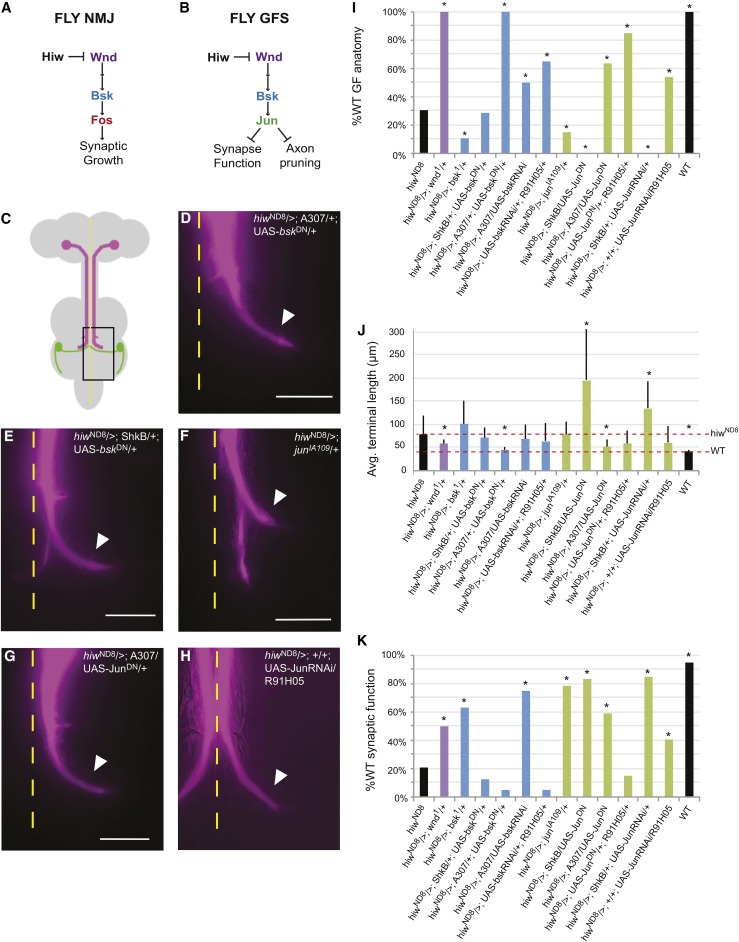
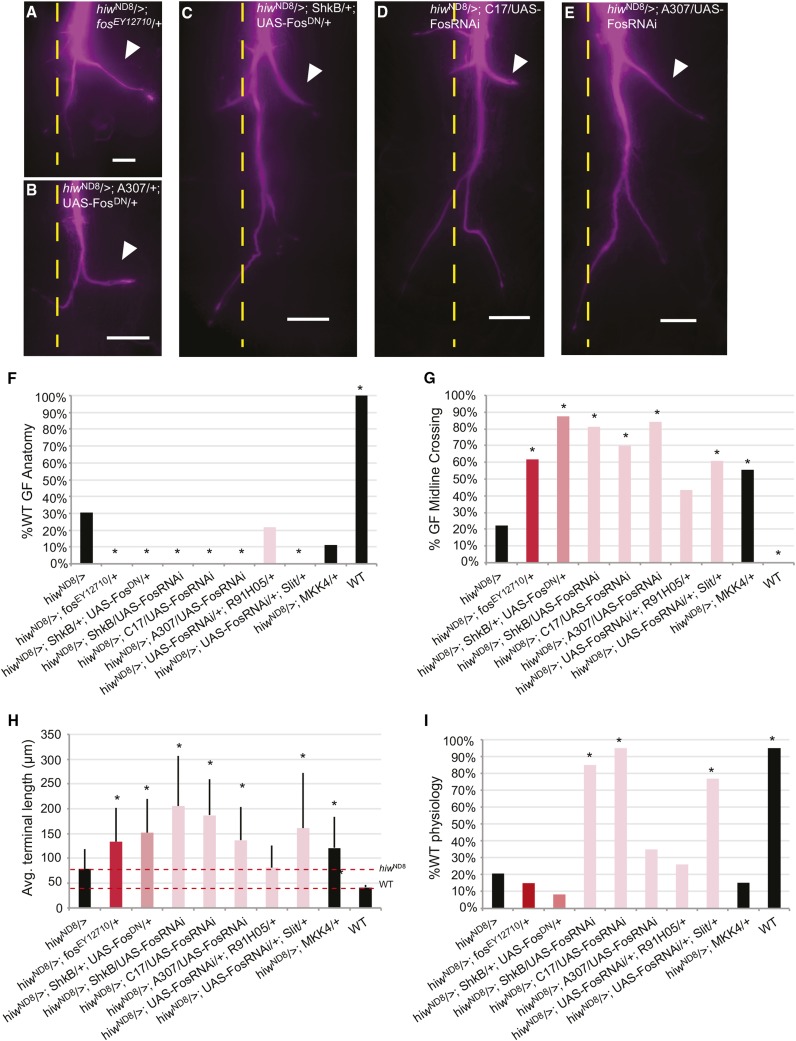
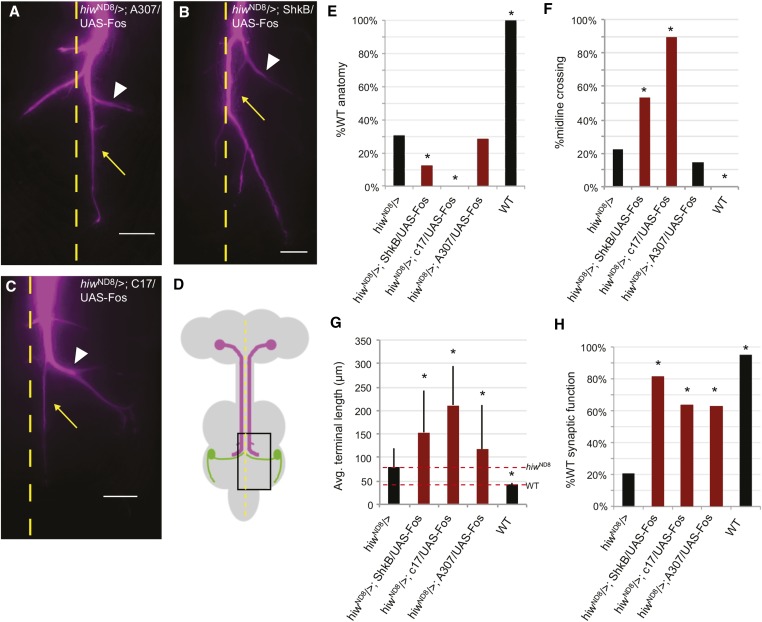
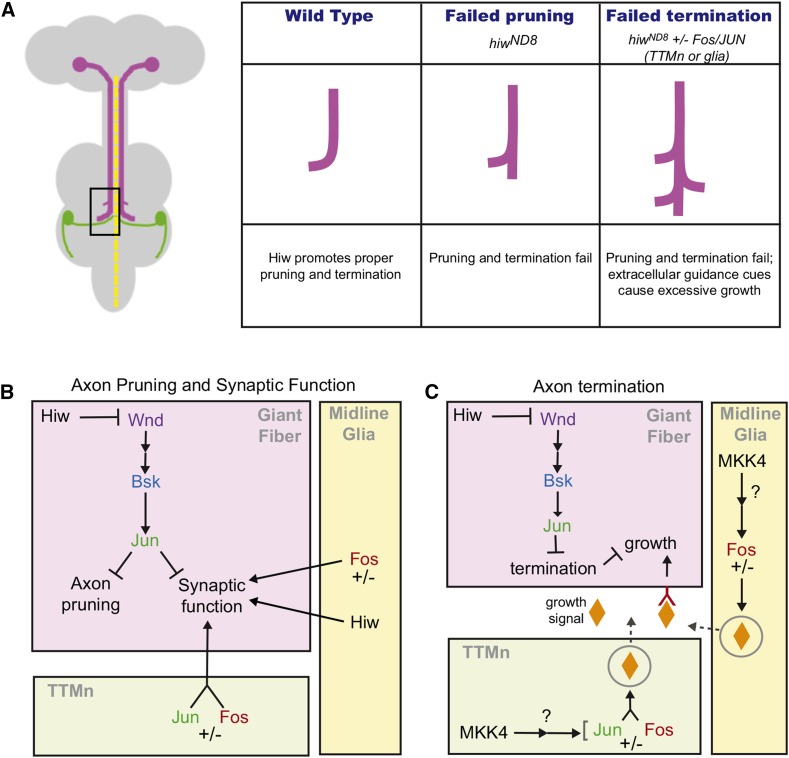
The ubiquitin ligase Highwire has a conserved role in synapse formation. Here, we show that Highwire coordinates several facets of central synapse formation in the Drosophila melanogaster giant fiber system, including axon termination, axon pruning, and synaptic function. Despite the similarities to the fly neuromuscular junction, the role of Highwire and the underlying signaling pathways are distinct in the fly's giant fiber system. During development, branching of the giant fiber presynaptic terminal occurs and, normally, the transient branches are pruned away. However, in highwire mutants these ectopic branches persist, indicating that Highwire promotes axon pruning. highwire mutants also exhibit defects in synaptic function. Highwire promotes axon pruning and synaptic function cell-autonomously by attenuating a mitogen-activated protein kinase pathway including Wallenda, c-Jun N-terminal kinase/Basket, and the transcription factor Jun. We also show a novel role for Highwire in non-cell autonomous promotion of synaptic function from the midline glia. Highwire also regulates axon termination in the giant fibers, as highwire mutant axons exhibit severe overgrowth beyond the pruning defect. This excessive axon growth is increased by manipulating Fos expression in the cells surrounding the giant fiber terminal, suggesting that Fos regulates a trans-synaptic signal that promotes giant fiber axon growth.
Keywords: Fos; giant fiber; highwire; pruning; synaptogenesis.
Copyright © 2017 by the Genetics Society of America.
Figures
References
-
- Allen M. J., Drummond J. A., Moffat K. G., 1998. Development of the giant fiber neuron of Drosophila melanogaster. J. Comp. Neurol. 397: 519–531. - PubMed
-
- Atwood H. L., Govind C. K., Wu C. F., 1993. Differential ultrastructure of synaptic terminals on ventral longitudinal abdominal muscles in Drosophila larvae. J. Neurobiol. 24: 1008–1024. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
