

Follicular CD8 T cells accumulate in HIV infection and can kill infected cells in vitro via bispecific antibodies
- PMID: 28100833
- PMCID: PMC5497679
- DOI: 10.1126/scitranslmed.aag2285
Follicular CD8 T cells accumulate in HIV infection and can kill infected cells in vitro via bispecific antibodies
Abstract
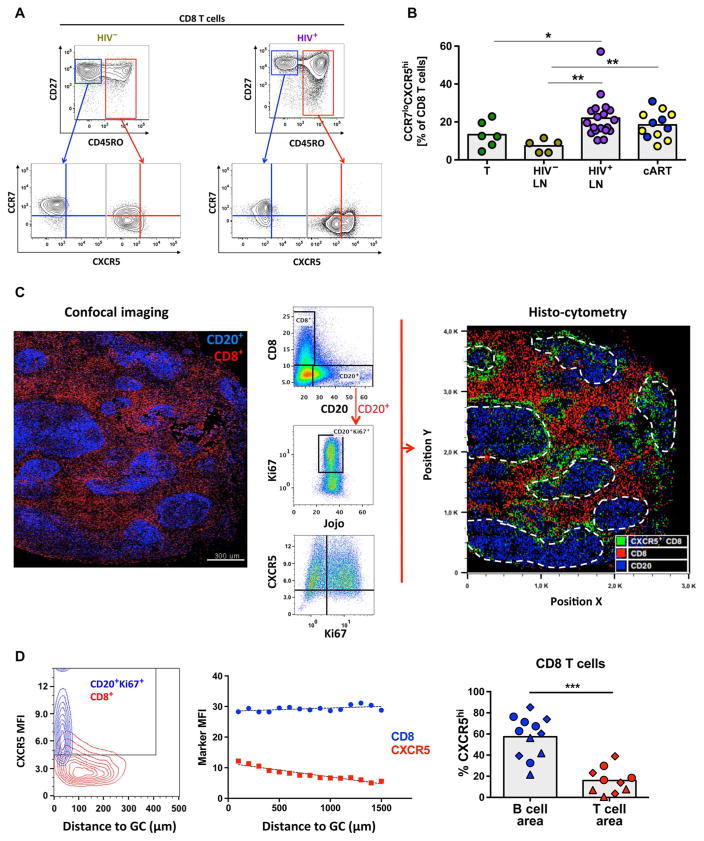
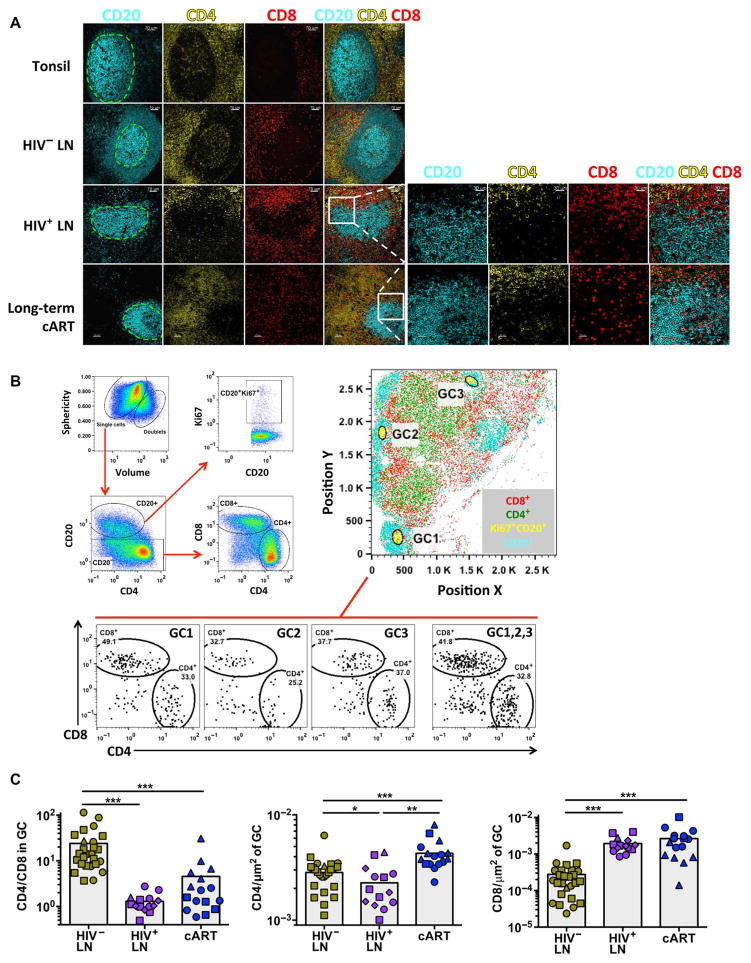
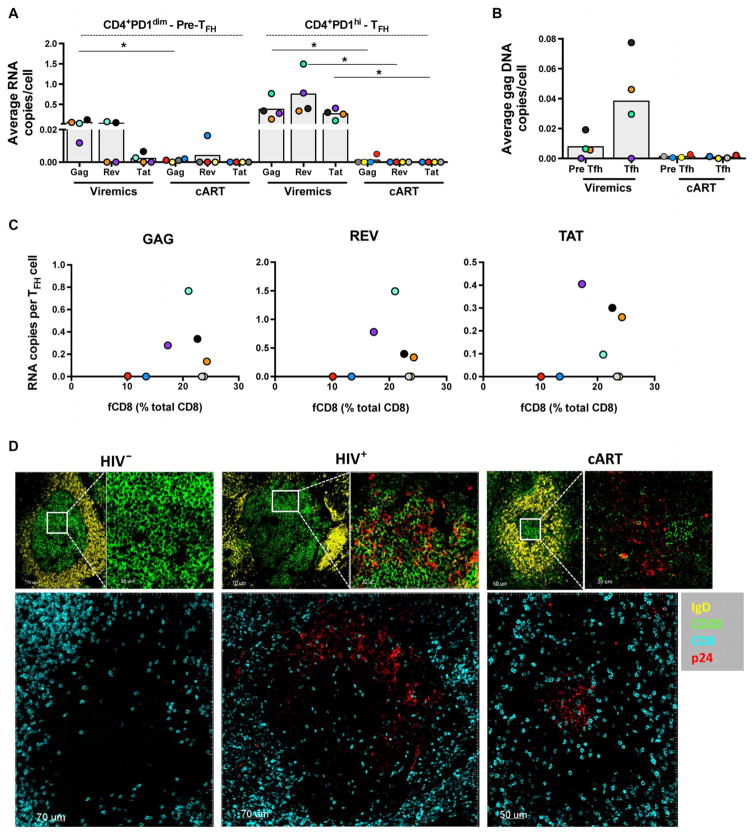
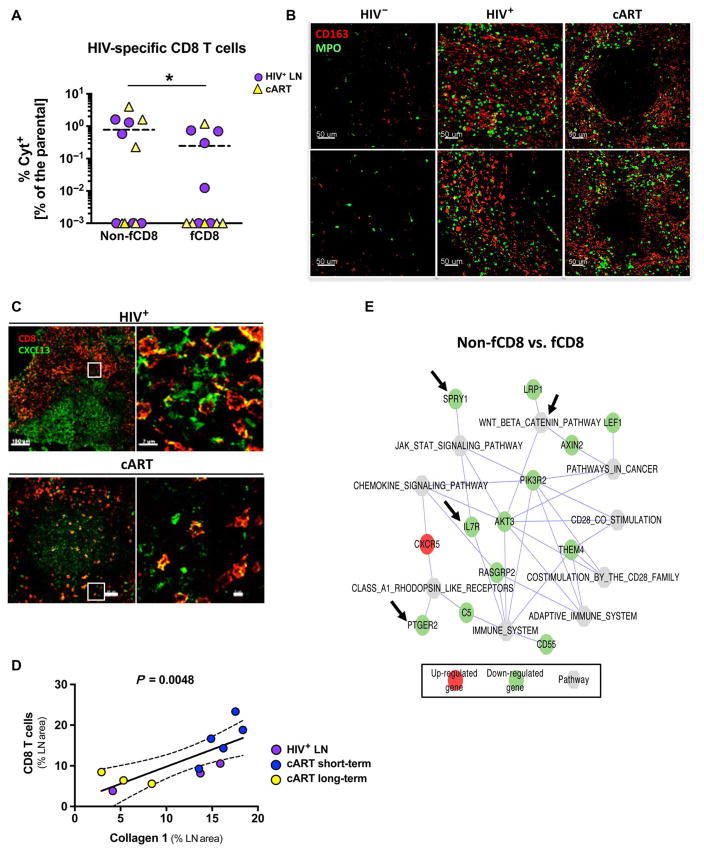
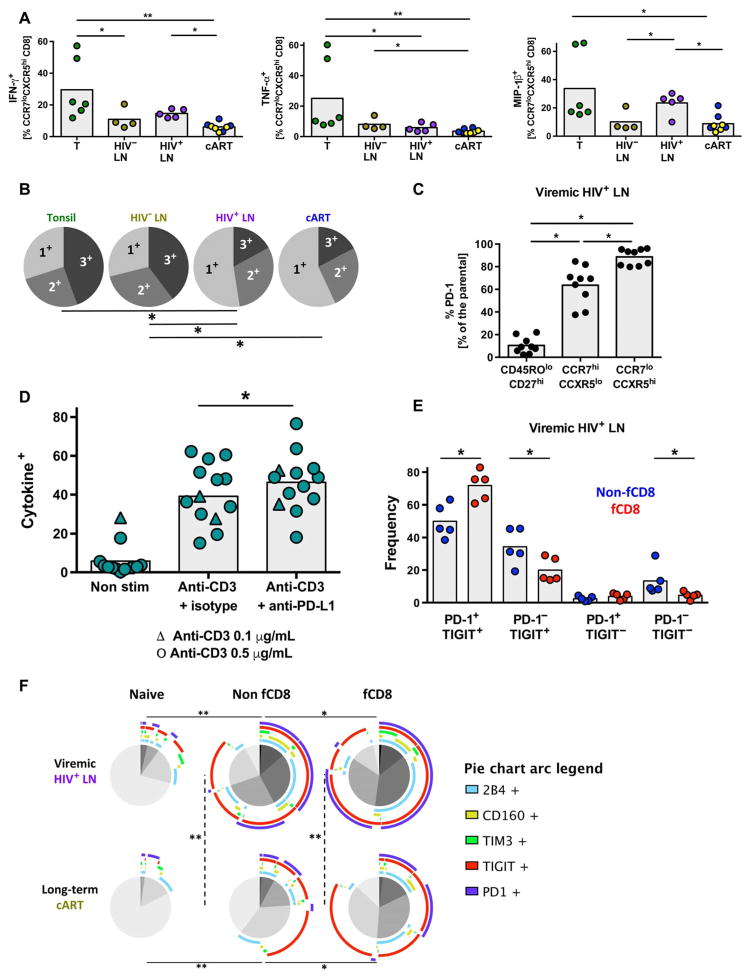
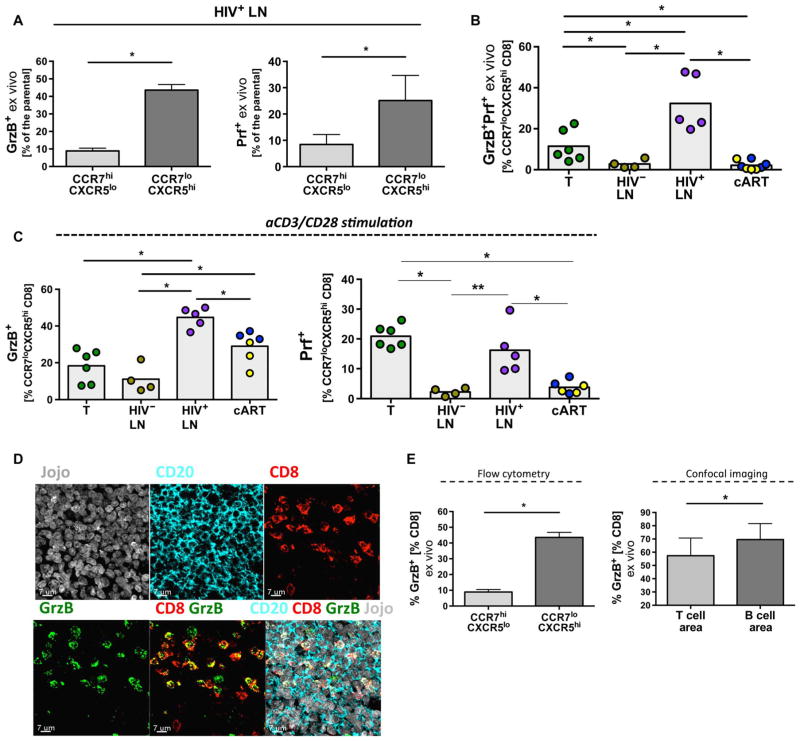
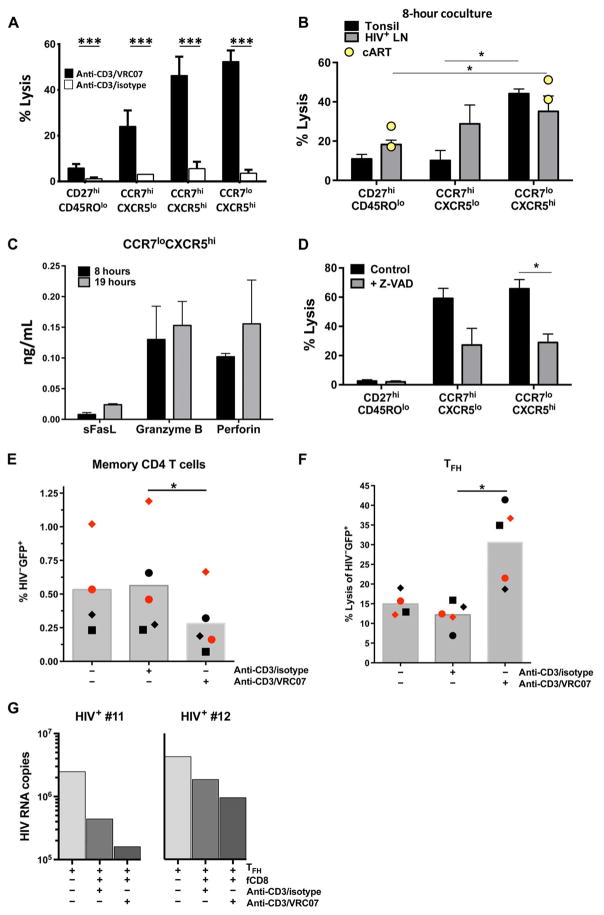
Cytolytic CD8 T cells play a crucial role in the control and elimination of virus-infected cells and are a major focus of HIV cure efforts. However, it has been shown that HIV-specific CD8 T cells are infrequently found within germinal centers (GCs), a predominant site of active and latent HIV infection. We demonstrate that HIV infection induces marked changes in the phenotype, frequency, and localization of CD8 T cells within the lymph node (LN). Significantly increased frequencies of CD8 T cells in the B cell follicles and GCs were found in LNs from treated and untreated HIV-infected individuals. This profile was associated with persistent local immune activation but did not appear to be directly related to local viral replication. Follicular CD8 (fCD8) T cells, despite compromised cytokine polyfunctionality, showed good cytolytic potential characterized by high ex vivo expression of granzyme B and perforin. We used an anti-HIV/anti-CD3 bispecific antibody in a redirected killing assay and found that fCD8 T cells had better killing activity than did non-fCD8 T cells. Our results indicate that CD8 T cells with potent cytolytic activity are recruited to GCs during HIV infection and, if appropriately redirected to kill HIV-infected cells, could be an effective component of an HIV cure strategy.
Copyright © 2017, American Association for the Advancement of Science.
Conflict of interest statement
Figures
References
-
- Connick E, Mattila T, Folkvord JM, Schlichtemeier R, Meditz AL, Ray MG, McCarter MD, MaWhinney S, Hage A, White C, Skinner PJ. CTL fail to accumulate at sites of HIV-1 replication in lymphoid tissue. J Immunol. 2007;178:6975–6983. - PubMed
-
- Embretson J, Zupancic M, Ribas JL, Burke A, Racz P, Tenner-Racz K, Haase AT. Massive covert infection of helper T lymphocytes and macrophages by HIV during the incubation period of AIDS. Nature. 1993;362:359–362. - PubMed
-
- Folkvord JM, Armon C, Connick E. Lymphoid follicles are sites of heightened human immunodeficiency virus type 1 (HIV-1) replication and reduced antiretroviral effector mechanisms. AIDS Res Hum Retroviruses. 2005;21:363–370. - PubMed
-
- Hufert FT, van Lunzen J, Janossy G, Bertram S, Schmitz J, Haller O, Racz P, von Laer D. Germinal centre CD4+ T cells are an important site of HIV replication in vivo. AIDS. 1997;11:849–857. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
